|
|
Компоненты лишайника и их взаимоотношения12 В одних источниках литературы лишайники определяют как очень интересную и своеобразную группу низших растений. (Горленко М.В., 1978). В других лишайники рассматривают как грибы, включающие в свое тело водоросли, — так называемые лихенизированные грибы, которые перешли к паразитизму на водорослях и в связи с этим значительно изменились. При таком подходе виды лишайников распределяются среди грибов в соответствии с типом плодового тела лишайникового гриба и особенностями его строения. Однако своеобразная форма тела лишайников, присутствие особых веществ, неизвестных ни у грибов, ни у водорослей, особенности обмена веществ и ряд других признаков позволяют рассматривать лишайники как самостоятельную группу организмов. (Горленко М.В., 1978). В лишайнике сочетаются два организма с противоположными свойствами: водоросль, которая составляет фикобионт и в процессе фотосинтеза создает органическое вещество, и гриб, который составляет микобионт, потребляющий это вещество. Тело лишайника представляет собой слоевище – таллом, причём основная часть объёма слоевища (90 – 95%) приходится на гифы гриба. Микобионт обычно представлен сумчатыми (Ascomycetes) или, гораздо реже, базидиальными (Basidiomycetes) грибами. Строение гиф в лишайнике имеет ряд особенностей. Гифы членистые, причём отверстия, соединяющие соседние компартменты, имеются не только в поперечных, но и в продольных перегородках. Для уменьшения потери воды клеточные стенки гиф сильно утолщены, особенно у гиф, расположенных во внешнем слое. Поскольку поперечные перегородки, разделяющие отсеки гифы, также утолщены, в этих местах увеличивается и диаметр. Для взаимодействия водорослей и правильной их ориентации в теле лишайника имеются особые двигающие гифы, которые могут перемещать клетки водоросли в нужное место (Приложение 1.). Другой особенностью микобионта является наличие ищущих и охватывающих гиф, которые необходимы для улавливания из внешней среды водорослей в процессе формирования таллома (Приложение 2.). В нижней части слоевища часто имеются жировые гифы, содержащие большое количество жира, интересно, что чаще всего такие гифы имеются у лишайников, живущих на известняках. Значение этих гиф ещё не выяснено. (Билич Г.Л., 2005). (Приложение 3.). В составе фикобионта лишайника встречаются сине-зелёные водоросли (по современным взглядам – это цианобактерии, которые относятся к надцарству прокариот) и различные представители настоящих (эукариотических) водорослей – зелёные, жёлтые и у одного вида из рода Веррукария (Verrucaria), живущего на калифорнийском побережье Тихого океана в качестве фикобионта обнаружена бурая водоросль Petroderma maculiforme (Пчёлкин А.В., 2006). Установлено, что для гриба не имеет особого значения, какого вида водоросль ему захватить, обычно его гифы стремятся захватить любые автотрофные клетки, находящиеся в пределах досягаемости. Однако далеко не все виды водорослей способны сосуществовать с грибом, большинство из них не выдерживает и погибает. Только самые выносливые и неприхотливые водоросли способны занять место фикобионта в лишайнике (Приложение 4). Лихенологии выяснили, что около половины из всех известных лишайников в качестве фикобионта имеют хлорококковую водоросль требуксию (Treboúxia). Из других зелёных водорослей в лишайниках встречается хлорелла (Chlorella), псевдохлорелла (Pseudochlorella), хлорококкум(Chlorococcum), а также улотриксовые (трентеполия (Trentepohlia), лептозира (Leptosira), фикопельтис (Phycopellis) и др.). Жёлто-зелёные водоросли в составе лишайник бывают очень редко, известны лишь два вида, содержащие гетерококкус (Heterococcus). Бурая водоросль (петродерма (Petroderma) найдена только у одного вида лишайников. Достаточно часто фикобионтом являются сине-зелёные водоросли (носток (Nostoc), глеокапса (Gloeocapsa), хроококкус (Chlorococcus) и др.) (Билич Г.Л., 2005). Находящиеся в составе лишайника водоросли тоже изменяют свою морфологию и зачастую их трудно узнать, сравнивая со свободными особями. Резко замедляется рост, отсутствие запасных трофических включений, поскольку значительная часть синтезированных органических веществ поглощается грибом. Однако сожительство с грибом закаляет водоросль, она способна выдерживать высушивание, высокие температуры. Но, в отличие от микобионта, водоросли, образующие фикобионт, вовсе не так заинтересованы в сожительстве с грибом, они прекрасно выживают и в свободном состоянии. Традиционно взаимоотношения микобионта и фикобионта определяются как взаимовыгодные, т.е. симбиотические, при которых гриб защищает водоросль от высыхания, нагревания, избыточных солнечных лучей и т.д., а также снабжает её неорганическими веществами, в том числе и водой. Водоросль, в свою очередь, снабжает оба компонента синтезированными органическими веществами. Между тем их взаимоотношения значительно сложнее. В природе не принято добровольно что-то отдавать представителям других видов, чаще всего имеет место обычный отъём, в том числе и нужных веществ. Ещё С. Швенденер, открывший двойственную природу лишайников, выдвинул гипотезу о паразитизмегриба на водоросли. В 1873 г. французский учёный Е. Борне обнаружил, что гифы гриба образуют боковые выросты – гаустории, проникающие в клетки водоросли, через которые отбирают необходимые вещества, т.е. проявляют признаки настоящего паразитизма (Билич Г.Л., 2005). Применение новейших методов физиологических исследований с использованием меченых атомов углерода и азота, особых красящих веществ и некоторых других позволило установить, что гриб получает вещества, ассимилируемые водорослью, и ведет себя в слоевище лишайника как паразитический организм (http://zoo.rin.ru/cgi-bin/index. 24.04.12). Дальнейшее исследование анатомии лишайников показало, что гриб способен формировать несколько типов всасывающих структур (Приложение 5). Ггаустории могут быть двух типов: интрацеллюлярные, если они глубоко проникают внутрь протопласта водорослевой клетки, и интрамембранные, которые только прорывают оболочку клетки, но не углубляются далеко в протопласт. Наряду с различными гаусториями гриб может образовывать другой тип боковых выростов – импрессории,которые вообще не разрушают клеточную оболочку водоросли, а только вдавливают её. Особенно часто и в больших количествах импрессории образуются у лишайников, обитающих в сухих местах. Третий тип всасывающей структуры – аппрессория– представляет собой не боковой вырост, как предыдущие типы, а концевую часть гифы, которая упирается в клеточную стенку водоросли, плотно к ней прижимается, но не повреждает её и не вдавливает в протопласт. Часто специализированные структуры не образуются, а необходимые грибу вещества он получает посредством тонкостенных обволакивающих гиф, которые оплетают клетки водорослей, но оставляют интактными оболочки. Если водоросль имеет нитчатую структуру, то она может быть оплетена слившимися гифами, образующими вокруг водоросли полую трубку. Таким образом, видно, что гриб, судя по всему, ведёт себя как паразит, отбирая у автотрофного организма синтезируемые им вещества. Однако для существования как самого гриба, так и лишайника в целом необходимо, чтобы водоросль, окруженная со всех сторон грибными гифами, все-таки могла жить и более или менее нормально развиваться. Если гриб начнет проявлять себя слишком активно, поражать все без исключения водоросли и, использовав их содержимое, уничтожать их, это, в конце концов, может при вести к гибели всех водорослей слоевища. Но тогда, уничтожив весь свой запас питания, погибнет и сам гриб, а значит, перестанет существовать и лишайник. Гриб должен использовать лишь часть водорослей, оставляя резерв - здоровые и нормальные водоросли, содержимым которых он мог бы питаться. Отношения между мико - и фикобионтом в слоевище лишайника не сводятся только к паразитизму гриба на водоросли. Ученые предполагают, что эти отношения гораздо сложнее. Следует отметить, что водоросль тоже «предпринимает» ответные действия для нейтрализации агрессии гриба. Например, у молодых клеток образуются более толстые оболочки, препятствующие развитию гаустории, поэтому грибом поражаются обычно более старые клетки, успевшие вырасти. Часто клетка, в которую внедрилась гаустория, немедленно приступает к делению, плоскость которого проходит непосредственно через участок с гаусторией. В результате дочерние клетки оказываются вне гаустории. Водоросль тоже проявляет потребность в определённых веществах, будучи автотрофным организмом, она способна самостоятельно синтезировать органические вещества. Однако окружённая гифами гриба водоросль не может поглощать воду и органические соли извне, поэтому ей приходится добывать их из тех же гиф. Следовательно, водоросль также проявляет признаки паразитизма. Хоть и в меньшей степени, чем гриб. Первым пришёл к выводу, что взаимоотношения гриба и водоросли представляют собой взаимный паразитизм, выдающийся отечественный лихенолог А.Н. Окснер (Билич Г.Л., 2005). Однако все высказанные по этому поводу точки зрения до сих пор остаются лишь гипотезами и большей частью не подтверждены экспериментально: лишайники оказались очень трудным объектом для физиологических исследований. Ученые пока не научились выращивать и поддерживать в живом состоянии слоевище лишайников в искусственных условиях. Время от времени появляются сообщения об удачных опытах выращивания лишайника в условиях лаборатории, но пока эти сообщения единичны и не всегда достоверны (http://zoo.rin.ru/cgi-bin/index.pl?idr=705&art=787 24.04.12). Одной из причин неудач подобных попыток можно считать чрезвычайно медленный рост лишайников. Обычно возраст взрослых слоевищ, которые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20 - 50 лет. В северных тундрах возраст некоторых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, имеющих вид корочки, в год дает прирост всего 0,2 - 0,3 мм. Описанные выше формы контакта между гифами микобионта и клетками водорослей, по всей видимости, не исчерпывают всего многообразия способов, с помощью которых гриб и водоросль в слоевище лишайников устанавливают между собой тесную связь. Исследования в этом направлении только начинаются. Таким образом, на основании специфических свойств лишайников можно дать следующее определение: лишайниками являются организмы, тело (слоевище) которых всегда состоит из двух компонентов - автотрофного фикобионта и гетеротрофного микобионта, образующих единое симбиотическое сожительство, отличающееся свободными морфологическими типами и особыми физиолого-биохимическими процессами. (http://textreferat.com.ua/referat2.php?id=3422&p=2 6. 05. 12). Взаимоотношения гриба (микобионта) и водоросли (фикобионта) в лишайнике настолько глубоки, что появляются совершенно новые морфологические формы, совершенно не похожие ни на грибы, ни на водоросли. Традиционно считается, что взаимоотношения гриба и лишайника симбиотические, однако многие выдающиеся учёные доказали, что гриб проявляет признаки паразитизма. Гифы гриба образуют боковые выросты, проникающие в клетки водоросли, через которые отбирают необходимые вещества. А.Н. Окснер первый выдвинул гипотезу о взаимном паразитизме, доказав что и водоросль проявляет признаки паразитизма, хотя и в меньшей степени, чем гриб. Микобионт и фикобионт в составе лишайника демонстрируют чрезвычайно сложные и противоречивые взаимоотношения. Морфология лишайников Тело лишайников не дифференцировано на органы и представляет собой слоевище (таллом). Слоевище лишайников очень разнообразно по окраске, размерам, форме и строению. Часто лишайники окрашены в различные цвета из-за присутствия пигментов (фиолетовые, синие, зелёные, коричневые, красные), которые откладываются в оболочке или цитоплазме гиф. Наиболее богаты пигментами гифы корового слоя лишайников и различные части их плодовых тел. У лишайников различают пять групп пигментов: зеленые, синие, фиолетовые, красные, коричневые. Механизм образования их до сих пор не выяснен, но совершенно очевидно, что важнейшим фактором, влияющим на этот процесс, является свет (О http://dic.academic.ru/dic.nsf/enc_biology/1445/%D0%9C%D0%BE%D1%80%D1%84%D0%BE%D0%BB%D0%BE%D0%B3%D0%B8%D1%8F#sel=27.04.12). Окраску слоевища также могут определять свойственные только этим организмам лишайниковые кислоты (они откладываются в виде кристаллов на поверхности гиф). Большинство лишайниковых кислот бесцветны, но некоторые из них окрашены, и иногда очень ярко — в желтый, оранжевый, красный и другие цвета. Окраска кристаллов этих веществ определяет и окраску всего слоевища. И здесь важнейшим фактором, способствующим образованию лишайниковых веществ, является свет. Чем ярче освещение в месте произрастания лишайника, тем ярче он окрашен. Как правило, очень ярко окрашены лишайники высокогорий и полярных районов Арктики и Антарктики. Это тоже связано с условиями освещения. Для высокогорных и полярных районов земного шара характерны большая прозрачность атмосферы и высокая интенсивность прямой солнечной радиации, обеспечивающие здесь значительную яркость освещения. В таких условиях в наружных слоях слоевищ концентрируется большое количество пигментов и лишайниковых кислот, обусловливая яркую окраску лишайников. Предполагают, что окрашенные наружные слои защищают нижележащие клетки водорослей от чрезмерной интенсивности освещения http://dic.academic.ru/dic.nsf/enc_biology/1445/%D0%9C%D0%BE%D1%80%D1%84%D0%BE%D0%BB%D0%BE%D0%B3%D0%B8%D1%8F#sel=27.04.12 Лишайники, обитающие в холодном климате, часто окрашены в тёмные цвета или даже становятся чёрными. Это тоже объясняется высокой концентрацией зеленых, синих и фиолетовых пигментов в коровом слое и плодовых телах этих лишайников. Хорошо пигментированные наружные слои антарктических лишайников не только защищают клетки водорослей от чрезмерно интенсивного освещения, в данном случае темная окраска слоевища является приспособительной и служит для притягивания тепловых лучей. По внешнему виду различают три типа талломов лишайников: накипные (или корковые), листоватые и кустистые; эти типы связаны между собой переходными формами. Кроме них не редко различают ещё чашуйчатый и филаментозный (нитевидный) типы талломов. (Горленко М.В., 1978). Накипные лишайники имеют вид тонкого налёта, или более толстой (несколько мм) корочки, прочно срастающейся с поверхностью субстрата. Их поперечные размеры обычно не большие (несколько см), но они могут сливаться образуя довольно крупные пятна. Самыми примитивными накипными лишайниками считаются лепрозные слоевища в виде тонкого порошкообразного налёта, состоящего из комочков водорослей, окружённых грибными гифами. Такие комочки легко отрываются и переносятся животными, ветром или водой, что обеспечивает их широкое распространение (рис. 1).
Часто таллом делится на отдельные фрагменты, разделённые трещинами, которые называются ареолами, а само слоевище – ареолированным. Такие слоевища образуются только на камнях, при этом ареолированная структура помогает выдерживать тепловое расширение скальной породы при нагревании. Чаще всего таллом накипного лишайника развивается на поверхности субстрата, однако некоторые виды частично или целиком погружаются в субстрат (Билич Г.Л., 2005).. Листоватые лишайники имеют вид уплощённой в дозовертральном направлении пластинки, чаще всего округлой формы. В составе таллома может иметься лишь одна пластинка, тогда он называется монофильным, или несколько – полифильные, причём пластинки могут быть цельными или рассечёнными. Обычно верхняя и нижняя поверхности слоевища имеют разную окраску. Таллом срастается с субстратом посредством ножки – гомфа, состоящего из грибных гиф. Такое прикрепление значительно менее прочно, чем у накипных лишайников, поэтому слоевище листоватых лишайников значительно легче отделяется от субстрата. Некоторые виды вообще не прикрепляются к субстрату, их слоевище свободно перекатываются ветром, т.е. они представляют собой кочующую форму (Рис. 2). Листоватые лишайники являются более высокоорганизованными, чем накипные. Таллом у них крупнее (10 – 20 см) и более дифференцирован. Переходной морфологической формой от накипных к листоватым лишайникам считаются чешуйчатые лишайники.
Рис. 2. Листоватые лишайники
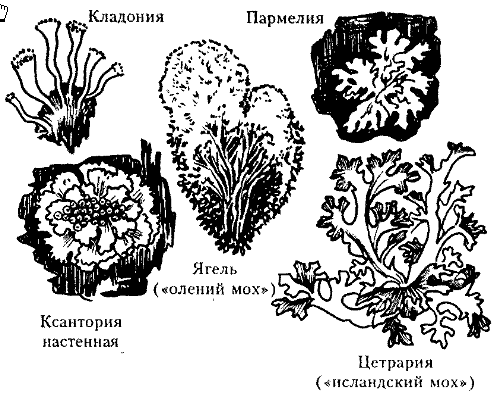
Кустистые лишайники являются самыми высокоорганизованными. Их слоевище имеет вид разветвлённого кустика, прикреплённого к субстрату посредством основания таллома – псевдогомфа. Значительно реже встречаются неветвящиеся формы в виде прямостоячего столбика. Ветвление кустистых лишайников возможно благодаря способности гиф расти не только в горизонтальном (как у накипных и листоватых), но и в вертикальном направлениях, образуя изгибы. Внешний вид кустистых лишайников может быть очень разнообразным. Кустистые лишайники демонстрируют исключительную жизнеспособность, их обнаруживали в Антарктиде на расстоянии всего 500 км от Южного полюса (Рис. 3).
Рис.3. Кустистые лишайники: 1 – цитрария, 2 – кладония, 3 – уснея Для некоторых кустистых лишайников характерен так называемый первичный таллом, накипный или чаще листоватый, состоящий из мелких чешуек. На первичном талломе уже развивается вторичный, собственно кустистый, таллом в виде отдельных неразветвленных или разветвленных, вертикально стоящих веточек. Эти ветви вторичного таллома называются подециями и особенно характерны для рода кладония (Cladonia), у которого они очень разнообразны по форме. Подеции могут быть палочковидные, шиловидные, туповатые вверху или расширенные в виде бокальчика (сцифовидные), в виде кустиков с обильно разветвленными ветвями. Сцифовидные подеции часто пролиферируют, т. е. из центра сцифы или по ее краям вырастают новые сцифовидные подеции. У лишайников из родов кладония и стереокаулон (Stereocaulori) на подециях развиваются чешуйки - филлокладии. Они соответствуют по происхождению чешуйкам первичного таллома, как бы переходящим на подеции. Первичный таллом может сохраняться в течение всей жизни лишайника, но у ряда видов он быстро исчезает и остаются только подеции. Талломы большинства кустистых лишайников имеют радиальное строение (у кладонии, стереокаулона, уснеи, алектории). Есть кустистые лишайники с лентовидным ветвящимся талломом, где морфологически хорошо различаются две стороны - верхняя и нижняя (так называемое дорзовентральное строение). Такой таллом характерен для рода цетрария (Celraria) (Горленко М.В.,1978). Таллом чешуйчатых лишайников представляет из себя небольшого размера чешуйки, иногда приподнимающиеся над субстратом одной из своих сторон (http://www.ecosystema.ru/07referats/pchelkin/poplich03.htm26.04.12). Таким образом, тело лишайников не дифференцировано на органы и представляет собой слоевище (таллом). По внешнему виду различают накипные, листоватые и кустистые лишайники, отличающиеся также между собой по размерам и строению. В зависимости от освещённости, а также от наличия лишайниковых кислот окраска лишайников может быть самой разнообразной. Анатомия лишайников Большую часть таллома лишайника занимают гифы гриба, относительный объём водорослевых клеток значительно меньше. В зависимости от взаимного расположения гриба и водоросли различают гомеомерные и гетеромерные слоевища. В первом случае водоросли распределяются среди грибных гиф без особого порядка. Такая более примитивная организация таллома характерна для слизистых лишайников. фикобионтом в них обычно являются сине-зелёные водоросли. Слоевища слизистых лишайников в сухом состоянии имеют вид темных или даже черных твердых и хрупких корок или пленок. Но они обладают одной интересной особенностью — способностью впитывать огромное количество воды, в 20 — 30 раз превышающее их собственную массу. При увлажнении они сильно разбухают, ослизняются, приобретают характер студня или желе и становятся оливково-зелеными или грязно-зелеными. Примером может служить влажное слоевище слизистого лишайника лептогиума. (Тахтаджян А.Л., 1974). Значительно чаще встречается гетеромернаяструктура, при котором слоевище дифференцировано на функциональные слои. При этом, чем сложнее морфологическое строение слоевища лишайника, тем сложнее и его анатомическая структура. У этих растений в ходе эволюции усложнение морфологического строения слоевища шло параллельно усложнению его анатомической структуры. У накипных лишайников различаются расположенный снаружи коровой слой, под ним находится слой водорослей и наиболее близко к субстрату – сердцевина. У большинства листоватых лишайников, более эволюционно продвинутых форм, в связи с отделением от поверхности субстрата происходит образование еще одного корового слоя — с нижней стороны слоевища. В таких слоевищах можно различить уже четыре слоя: верхний коровой слой, слой водорослей, сердцевину и нижний коровой слой. У кустистых лишайников с плоскими лентовидными лопастями образуется еще один анатомический слой — слой водорослей с нижней стороны слоевища. У этих лишайников в слоевище можно различить уже пять анатомических слоев: два коровых слоя на верхней и нижней поверхности слоевища, два слоя водорослей, тоже с верхней и нижней стороны лопастей, и сердцевину. И наконец, кустистые лишайники с округлыми лопастями, обладающие наиболее высокоорганизованными слоевищами, имеют радиальную структуру: снаружи лопасти этих лишайников покрыты коровым слоем, под ним расположен слой водорослей, а центральная часть слоевища заполнена сердцевиной (Приложение 6). Каждый из перечисленных анатомических слоев слоевища выполняет в жизни лишайника ту или иную функцию и в зависимости от этого имеет совершенно определенное строение. Коровой слой играет в жизни лишайника очень важную роль. Он выполняет сразу две функции: защитную и укрепляющую. Он защищает внутренние слои слоевища от воздействия внешней среды, прежде всего водоросли от чрезмерного освещения. Поэтому коровой слой лишайников обычно бывает плотного строения и окрашен в сероватый, коричневый, оливковый, желтый, оранжевый или красноватый цвет. Коровой слой служит и для укрепления слоевища. Чем выше слоевище поднимается над субстратом, тем более оно нуждается в укреплении. Укрепляющие механические функции в таких случаях нередко выполняет толстый коровой слой. Гифы, образующие коровой слой, могут тесно переплетаться, формируя внешнее подобие паренхиматозной ткани. Однако настоящая тканевая организация формируется при трёхмерном делении клеток, здесь же этого не происходит (гифы нарастают своими концами), поэтому структурные ансамбли корового слоя лишайника называются параплектенхимой, реже псевдопаренхимой ( Рис. 4).
Рис.4. Параплектенхимный коровой слой (Peltigera) (а – коровой слой, б – клетки водоросли, в – сердцевинные гифы)(по А. Л. Тахтаджяну)
У многих лишайников в коровом слое оболочки клеток гиф бывают сильно утолщены, а просветы клеток, заполненные протоплазмой, очень узкими. При срастании таких гиф образуется коровой слой, представляющий сплошную хрящевидную массу, где границы клеток незаметны, а их просветы кажутся почти точками. При этом гифы на своей поверхности выделяют желатин, плотно цементирующий их между собой. Такого типа ткани у лишайников называют прозонлектенхиматическими (Тахтаджян А.Л., 1974). Плотный хрящевидный коровой слой этих лишайников не только надежно защищает нижележащие слои водорослей, но и хорошо укрепляет приподнимающееся над землей слоевище Реже гифы корового слоя лишайников не срастаются, а лежат свободно параллельно друг другу. При этом они могут образовывать два различных типа корового слоя. Если гифы расположены параллельными рядами перпендикулярно поверхности слоевища, то образуется палисадный коровой слой. Во втором случае, гифы корового слоя лежат не перпендикулярно, а параллельно поверхности слоевища и имеют вид удлиненных волокон. Коровой слой такого строения носит название волокнистого. На нижнем коровом слое лишайников обычно образуются органы прикрепления. Иногда они имеют вид очень тонких нитей, состоящих из одного ряда клеток. Эти нити называют ризоидами. Каждая такая нить берет начало от одной клетки нижнего корового слоя. Нередко несколько ризоидов соединяются в толстые ризоидальные тяжи. Их можно видеть, например, на нижней стороне крупнолистоватых слоевищ пельтигер. У листоватых лишайников рода пармелия образуются более толстые прикрепительные тяжи, называемые ризинами. В образовании ризин принимают участие не только гифы нижнего корового слоя, но и сердцевины. Снаружи они покрыты коровым слоем, а внутри образованы сердцевинными гифами. На самом конце ризины клетки гиф вытягиваются и расходятся в разные стороны, образуя кисточку. Нередко на конце такой кисточки образуется слизистая капелька, с помощью которой слоевище более плотно прикрепляется к субстрату. Иногда на конце ризины образуется особая прикрепительная пластиночка, которой лишайник, как маленькой подошвой, прикрепляется к поверхности коры дерева или камня. У некоторых листоватых лишайников, имеющих вид более или менее округлой листовидной пластинки, слоевище прикрепляется лишь в своей центральной части с помощью короткой ножки — гомфа или псевдогомфа. Таким образом, в зависимости от взаимного расположения гриба и водоросли различают гомеомерные,как наиболее примитивные, и гетеромерные слоевища. Гетеромерные слоевища делятся на слои (коровой слой, слой водорослей, сердцевина) и в зависимости от внешнего вида у разных лишайников выделяются разное количество слоёв. Так, у накипных лишайников выделяется три слоя, у листоватых – четыре, у кустистых – пять. Каждый из перечисленных анатомических слоев слоевища выполняет в жизни лишайника ту или иную функцию и в зависимости от этого имеет совершенно определенное строение. Размножение лишайников Размножение лишайников столь же своеобразно, как и вся их организация. Каждый из компонентов таллома способен размножаться индивидуально, например клетки водоросли время от времени делятся, однако последующее развитие нового таллома возможно лишь в случае повторной встречи гриба и водоросли. Поэтому более перспективным будет такой способ размножения, при котором генеративные структуры содержат оба компонента. Большинство лишайников размножается преимущественно вегетативным путём, причём для многих из них этот способ размножения остаётся единственным. Высыхая, слоевище становится очень хрупким и от него легко отламываются кусочки, которые затем подхватываются ветром и распространяются. Так происходит у многих лишайников тундры. Соредии и изидии являются специализированными структурами вегетативного размножения, причём каждая из них содержит элементы микобионта и фикобионта. Соредии (греч. soros – куча) представляют собой микроскопические комочки клеток водоросли, окружённые гифами гриба. Скопление соредий (они называются соралями) напоминают порошкообразную массу, которая покрывает сверху слоевище или окружает таллом по внешнему периметру. Соредии образуются в слоевище под верхней корой, в зоне водорослей. У разных лишайников процесс образования соредии происходит неодинаково, однако в общих чертах он всегда сводится к следующему. На месте образования соредий каждая клетка водорослей делится на несколько дочерних клеток. В то же время гифы гриба начинают сильно ветвиться, делятся на множество клеток и окружают каждую из дочерних клеток водоросли. В результате этого процесса под коровым слоем слоевища образуется масса водорослей, оплетенная коротковетвящимися многоклеточными гифами. Спустя некоторое время она распадается на отдельные клубочки, каждый из которых представляет собой соредий. Когда клубочков образуется очень много, их масса начинает давить на коровой слой лишайника, тот не выдерживает, разрывается и соредий выступают наружу, на поверхность слоевища. Здесь они образуют порошистую массу, которая легко сдувается и разносится ветром. Попавший в благоприятные условия соредий начинает разрастаться и сразу развивается в новое слоевище лишайника (Тахтаджян А.Л.,1974). Соредиями размножается главным образом высокоорганизованные лишайники (листоватые и кустистые), которые обычно не образуют плодовых тел. Обычно на поверхности слоевища соредий образуют скопления определенной формы, называемые соралями (Приложение 7). Изидии также свойственны листоватым и кустистым формам, но они образуются реже, чем соредии. Морфологически изидии представляют собой мелкие выросты на верхней поверхности таллома, внутри которых находятся водорослевые клетки, окружённые гифами. Снаружи изидии покрыты коровым слоем. Внешний вид изидии может быть самым разнообразным, но для каждого вида лишайников специфична определённая форма. Оторванные от слоевища изидии в благоприятных условиях разрастаются в новые слоевища. Однако значение изидий в размножении лишайников не так велико по сравнению с соредиями. Очевидно, это связано с тем, что обычно изидии не отрываются от слоевища так легко, как соредий, а обламываются лишь животными или человеком. Изидии не только служат для размножения, но и выполняют определенную роль в увеличении ассимиляционной поверхности слоевища лишайника (Приложение 8). Преимущество размножения лишайников соредиями и изидиями перед половым способом состоит в том, что в нем принимают участие оба компонента лишайника— и гриб, и водоросль. При половом размножении на слоевищах лишайников в результате полового процесса формируются половые спороношения в виде плодовых тел. В зависимости от систематического положения микобионта различают базидиальные и сумчатые лишайники. Базидиальные лишайники в качестве микобионта имеют базидиальные грибы. Это совсем небольшая группа (всего около 20 видов) имеет самостоятельное происхождение и распространена преимущественно в тропиках, хотя некоторые из них обитают в умеренном климате и даже в тундре. У небольшой группы лишайников споры образуются зкзогенно, на вершине удлиненно-булавовидных гиф — базидий. На верхушке базидий имеются четыре отростка — стеригмы, на концах которых развиваются четыре споры. Лишайники с таким образованием спор объединяются в группу базидиальных лишайников (Приложение 9). У сумчатых лишайников выделяют следующие основные типы плодовых тел: апотеции, гастротеции и перитеции.Большинство лишайников, свыше 250 родов, формируют открытые плодовые тела в виде апотециев — дисковидных образований. Эти округлые дисковидные образования, напоминающие по виду маленькие блюдца, очень часто можно наблюдать на верхней поверхности слоевища накипных и листоватых лишайников или на концах лопастей кустистых слоевища. Обычно их диаметр не превышает 1—2 мм, но у некоторых листоватых и кустистых лишайников он может достигать 1—3 см, а у накипных — всего 0,1—0,5 мм. По форме апотеции чаще всего округлые, реже овальные; когда же несколько апотециев развивается рядом друг с другом, они нередко становятся угловатыми. У семейства артониевых (Arthoniaceae) они имеют форму звезд или неправильных пятен. Обычно апотеции образуются на верхней поверхности слоевища лишайников, плотно прирастая к ней своей нижней частью или возвышаясь над ней на тонкой маленькой или довольно длинной ножке (рис.). У некоторых лишайников они бывают целиком погружены в слоевище, например у солорин. Только у двух родов лишайников — нефрома (Nephroma) и нефромопсис (Nephromopsis)— апотеции образуются не на верхней, а на нижней стороне лопастей.(Тахтаджян А.Л.,1974). В апотециях можно различить центральную часть — плоский диск и периферическую — округлый выпуклый валик, окружающий диск. Плодущей частью апотеция является диск. На его поверхности развивается плодущий слой, который носит название гимениального. Он образован вертикально стоящими сумками со спорами и расположенными между ними бесплодными гифами— парафизами. Парафизы — тонкие, нитевидные гифы (обычно толщиной 1,5—2 мкм), простые или разветвленные, со свободными верхними концами. Они служат для защиты сумок со спорами. По высоте они обычно несколько превосходят сумки, а их свободный верхний конец бывает булавовидно утолщен и окрашен в зеленоватый, синеватый, коричневатый, желтоватый и другие цвета. Эти окрашенные утолщенные вершинки, плотно прилегая друг к другу, образуют над сумками в верхней части гимениального слоя защитный слой, который называют эпитецием. От окраски эпитеция зависит окраска диска апотеции: он может быть коричневым, черным, желтым, оливковым, красным, оранжевым, розовым и др. Под гимениальным слоем расположен гипотеций— слой, состоящий из плотно переплетенных гиф. Тонкое цитологическое исследование позволяет различить в нем основания парафиз и аскогенный аппарат, из которого по мере созревания вырастают сумки. Край апотеция, имеющий вид округлого валика, как защитный футляр, одевает плодущий диск с боков и снизу. В зависимости от его строения различают несколько типов апотециев: леканоровые, лецидеевыеи биаторовые(Тахтаджян А.Л.,1974). Диск леканоровых апотециев окружен слоевищным краем, который всегда содержит внутри водоросли, а строение и окраска его очень похожи на строение и окраску слоевища. На анатомическом срезе в нем можно различить коровой слой, слой водорослей и слой рыхлой сердцевины. Леканоровые апотеции легко распознаются и по внешнему облику: окраска их края всегда отличается от окраски диска. Если диск может быть коричневым, черным, красным, то край чаще всего бывает более светлым, окрашенным в один цвет со слоевищем,— сероватым, беловатым, серовато-зеленоватым, светло-коричневым и т. д. (Приложение 10). Второй тип апотециев — лецидеевые апотеции — также легко можно распознать по внешнему облику. Обычно они черные и очень твердые, их с трудом можно раздавить на предметном стекле. В отличие от леканоровых край этих апотециев имеет ту же окраску, что и диск: он тоже черный и носит название собственного края. Собственный край никогда не содержит водорослей и отличается по строению от слоевища. У лецидеевых апотециев он состоит из темноокрашенных, плотно прилегающих друг к другу гиф, которые образуют с трех сторон вокруг диска валик, называемый эксципулом. (Приложение 11).
Биаторовыеапотеции по строению похожи на лецидеевые: у них тоже имеется собственный край, образованный эксципулом. Но, в отличие от лецидеевых апотециев, гифы, образующие эксципул, и сами биаторовые апотеции окрашены в светлые тона — они бывают восковидные, желтовато-коричневатые, красновато-коричневые, розовато - или беловато-желтые и никогда не бывают черные. Биаторовые апотеции отличаются очень мягкой консистенцией, их легко можно раздавить на предметном стекле, как теплый, расплавленный воск. Гастеротеции, как и апотеции, являются плодовыми телами открытого типа. В них также можно различить диск и край. Но они отличаются от апотециев сильно вытянутой, линейной формой. Обычно гастеротеции имеют вид штрихов, простых или разветвленных линий. Диск у этих плодовых тел очень узкий, напоминающий тоненькую полоску, в то время как край, который может быть' и собственным и слоевищным, сильно выступает и возвышается над глубоко погруженным диском. Гастеротеции у лишайников встречаются довольно редко. Они известны лишь у представителей двух родов накипных лишайников — графис(Graphis) иопеграфа(Opegrapha). 
12 Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
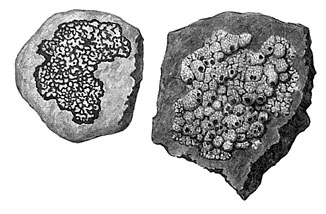 Рис. 1. Накипные лишайники (по Г.Л. Биличу)
Рис. 1. Накипные лишайники (по Г.Л. Биличу)