
|
|
Размножение и развитие папоротника.12
Летом на нижней поверхности листьев папоротника образуются группы бурых мешочков со спорами, покрытые особыми покрывальцами. В сухую жаркую погоду мешочки растрескиваются и споры при помощи ветра рассеиваются на дальние расстояния. Попав на влажную почву, они прорастают. Сначала развивается зеленая нить. Ее клетки делятся и образуют зеленую сердцевидную пластинку. Это — заросток папоротника. С нижней его стороны образуются ризоиды и половые органы. В мужском половом органе развиваются сперматозоиды, а в женском — яйцеклетка. Во влажную погоду многожгутиковые сперматозоиды проникают в женский орган, где и происходит оплодотворение. Образовавшаяся зигота делится, и из нее развивается молодой папоротник. Таким образом, папоротник, образующий споры, — это спорофит, а заросток папоротника — это гаметофит. В процессе жизни происходит смена двух поколений — спорофита и гаметофита.
7. По каким причинам мегафильный лист отличается от микрофильного? Как эти листы возникают? Для каких отделов характерны. Возникновение листьев в ходе эволюции шло двумя путями. Высшие растения лист как орган унаследовали после уплощения и срастания телломов – систем осей – ветвящегося вегетативного тела первичных наземных растений – риниофитов. Это представление развивает марфофильная теория эволюции и рассматривает лист как «плосковертку». Мегафильные листы характерны для Папоротников как споровых сосудистых растений. Лист покрыт кожицей с устьицами на нижней поверхности и пронизан жилками. В клетках основной ткани листа находятся хлоропласты, в которых осуществляется фотосинтез. В отличие от плаунов и хвощей для папоротниковидных характерна макрофилия(крупнолистность). У некоторых видов длина листьев может достигать 30 м и более. Современные ученые считают, что листья папоротников возникли в результате срастания и уплощения ветвей (теломов) древних предковых форм. Эту гипотезу подтверждает наличие у листьев апикальной (верхушечной) меристемы, что позволяет им расти верхушкой. У всех остальных растений верхушечный рост листа весьма непродолжителен, и листья растут основанием. Листья папоротников называют вайями. Молодые вайи свернуты в виде улитки и покрыты коричневыми чешуйками.
2. возникновение листа в результате поверхностных уплощенных выростов первичной коры на осевом органе – телломе, привел в образованию микрофильного листа и соответствует строению листа плаунов. Все побеги плауновидных довольно густо покрыты мелкими простыми листочками, форма которых может быть овальной, шиловидной, шиповидной, линейной, ланцетовидной. Располагаются такие листочки на побегах разных видов или поочередно, или супротивно, или мутовчато. Для этих плаунов характерна микрофилия, т. е. мелколистность. Листья на стебле обычно располагаются по спирали и подразделяются на вегетативные листья (трофофиллы) и спороносные листья (спорофиллы).
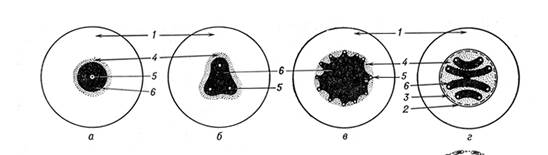
8. Начертите схему эволюции стелы у Плауновидных. Почему происходит разделение центрального цилиндра? Под стелой понимаем совокупность проводящих, механических и паренхимных тканей, ограниченных перициклом. Наиболее примитивным типом стелы является та, в которой проводящая ткань образует сплошную массу, и центральное ядро, состоящее из ксилемы, полностью окружено флоэмой. Такая стела называется протостелой. Протостела не только проста по структуре, но и представляет собой исходный тип, от которого в процессе эволюции произошли все другие типы стелы. Она была характерна для риниофитов и многих других вымерших форм. В современной флоре протостелу имеют некоторые плауны и папоротники. Наиболее примитивной формой протостелы является гаплостела (греч. гаплос - простой), состоящая из центрального пучка первичной ксилемы, окруженного цилиндром флоэмы. Более продвинутой формой является актиностела (греч. актинос - луч), имеющая радиальные выступы ксилемы в виде лучей (на поперечном срезе имеет вид звезды). При переходе к актиностеле флоэма и ксилема имеют большую поверхность соприкосновения с окружающими живыми тканями, что способствует лучшему проведению веществ. Типы стелы высших растений: 2 - протостела; б, в - актиностела; г - плектостела; 1 - первичная кора; 2 - эндодерма; 3 - перикл; 4 - флоэма; 5 - протоксилема; 6 - метаксилема; Исходным типом стелы, характерным для древнейших высших растений - псилофитов (риния), считают протостелу, имеющую вид центр, тяжа, во внутренней части которого расположена ксилема, окруженная нерезко отграниченной от первичной коры флоэмой. Совершенствование структуры стелы в эволюции растений, по мнению английского ботаника Ф. Боуэра, шло по пути создания наиболее рационального соотношения между объёмом и поверхностью проводящих тканей, что достигалось изменениями контуров стелы и привело к расчленению её на отдельные тяжи. В разных направлениях эволюции растений возникали различные типы структурной организации стелы. Так, изменения контуров ксилемы обусловили преобразование протостелы в актиностелу и плектостелу. Актиностела, характерная для псилофитов (астероксилон), а из современных растений - для псилота, имеет лопастные очертания поперечного сечения экзархной ксилемы. 9. В чем преимущества мегаспоры у разноспоровых растений в сравнении со спорами равноспоровых? Среди папоротникообразных существуют равноспоровые и разноспоровые растения. Большинство папоротников образует одинаковые по размерам и строению споры, т. е. являются равноспоровыми. Однако немногочисленная группа водных папоротников (сальвиниевые) – разноспоровые растения. У равноспоровых все споры одинаковые, из них образуются обоеполые заростки, несущие и антеридии, и архегонии. У разноспоровых растений в одних спорангиях развиваются микроспоры, в других – мегаспоры (макроспоры). Из микроспор вырастают редуцированные мужские заростки с антеридиями, из мегаспор – более крупные женские заростки с архегониями. Образующиеся в антеридии многочисленные дву- или многожгутиковые сперматозоиды выходят из него и проникают в архегоний, где один из них сливается с яйцеклеткой. Образуется диплоидная зигота. Оплодотворение у папоротникообразных (как и у моховидных) возможно лишь при наличии капельно-жидкой воды.
10. Можно ли утверждать, что мегаспорофилл Селагенелловых и семенная чешуя Голосеменныых являются гомологичными органами? Докажите. Гомологичными называются органы сходные по общему плану строения, положения и возникновения в процессе онтогенеза. Но разные по выполняемым функциям. Селагенелла относится к отделу плауновидных. Это наземное травянистое растение, высотой 8-12 см. стебли прямостоячие или лежачие. На верхушке некоторых побегов развиваются спороносные колоски. Листья спороносного колоска – спорофиллы – заметно отличаются от вегетативных. В пазухах спорофиллов на короткой ножке развивается мега – и микроспорангии. Спорофилы несут язычки, которые никогда не отваливаются. Важное отличие селагенелловых от плауновых – их разноспоровость: мегаспорангии содержат 4 мегаспоры, микроспорангии – множество спор. При разноспоровости наиболее радикальная редукция гаметофита (заростка) – гаплоида и развитие до гигантских размеров спорофита – диплоида. Разноспоровость вызвала раздельнополость, т.е. двудомность заростков. Рассмотрим размножение голосеменных на примере сосны обыкновенной. Порофиллы собраны в шишках 2-х видов: мужские представлены метельчатыми соцветиями, женские – одиночными. Мужская шишка элепсовидной формы она образуется в пазухе чешуйки, на месте укороченного побега и представляет собой побег с хорошо развитой осью, на котором спирально расположены микроспорофиллы – редуцированные спороносные листья (гомологи тычинок покрытосеменных). У основания оси имеются низовые листья в виде чешуек, играющих защитную функцию. Женские шишки развиваются на верхушках молодых побегов. На главной оси расположены мелкие бесплодные чешуйки, называющиеся кроющими. В пазухах этих чешуек образуются крупные толстые чешуи с двумя семязачатками на верхней стороне. Их называют семенными чешуями. Это редуцированные спороносные побеги, возникающие в пазухах кроющих чешуй. Женская шишка – это группа укороченных боковых побегов, находящихся на общей оси. Семязачатки находятся у основания семенной чешуи. Молодой семязачаток и есть мегаспорангий. Таким образом мегаспорофилл Селагенелловых и семенная чешуя голосеменных являются гомологами. Т.к. они сходные по общему плану строения, положения и возникновения в процессе онтогенеза, но выполняют разные функции. У селагенелловых – это образование мегаспор, у Голосеменных – семенная чешуя – место. где развиваются семязачатки.
11. Начертите схему воспроизведения селагенелловых и сальвинии. Укажите сходство и различие.
Рис. Сальвиния плавающая. А – спорофит; Б – лист. В – спорокарпии; Г – микроспорангий с мужским гаметофитом; Д -завершающие стадии развития мужского гаметофита с 8 сперматозоидами; Е – мегаспорангий; Ж- мегаспорангий с женским гаметофитом; З – архегогии; И – мегаспорангий с женским заростком и развивающимся зародышем спорофита; к – молодой спорофит.
Органы спороношения развиваются на нижней подводной стороне побега, на сегментах погруженных листьев. Спорангии находятся внутри спорокарпиев. Внешне они одинаковы по форме, размерам, но на одних содержатся мужские спорангии – микроспорангии, на других – женские – мегаспорангии. Стенка спорокарпия напоминает стенку листа сальвинии. Спорангии тонкостенные, мужские содержат по 64 микроспоры, женские – по 32 мегаспоры. Но лишь одна из них – центральная – жизнеспособна и может дать начало гаметофиту. При созревании спор спорокарпии опускаются на дно водоема. После их разрушения спорангии всплывают на поверхность. Микро- и мегаспоры, пророст ая образуют соответствующий гаметофит. Оба они прорастают внутри спорангиев. Мужской сильно редуцирован, женский виден невооруженным глазом. Архегоний с очень короткой шейкой. Глубоко погружен в ткань заростка. После полового процесса зигота дифференцируется на ножку, первый лист и верхушку стебля. У водяных папоротников наблюдается редукция гаметофита. Вегетативная часть мужского гаметофита представлена 2 клетками. Женский гаметофит менее редуцирован, но все же настолько мал, что значительная его часть помещена внутри мегаспоры. На первых фазах развития спорофит питается за счет зеленого женского гаметофита.
12. В чем смысл редукции заростков (гаметофитов) у разноспоровых Плауновидных и Папоротниковидных. Ответ поясните. В эволюции высших растений происходит постепенная редукция (уменьшение и упрощение) гаметофита и преобладание в жизненном цикле спорофита. Среди причин, которые обобусловилипреобладание спорофита, можно назвать его диплоидность. Она по сравнению с гаплоидностью гаметофита обеспечивает более высокий уровень синтетических процессов, а, с другой стороны, рецессивные мутации, снижающиежизнеспособность организма, при диплоидном состоянии не проявляютсяв фенотипе, то есть генотипически диплоидный организм можно считатьболее «стабильным». Спорофиты большинства отделов высших растений (плауновидных, хвощевидных, папоротниковидных) представляют собой крупные многоклеточные организмы со сложным анатомическим строением и расчленением тела на органы – стебли, листья, корни (настоящие или придаточные). Когда мы представляем себе растение плауна, хвоща, папоротника, мы обычно подразумеваем его спорофит. На спорофитах образуются органы спороношения – спорангии. У высших растений спорангий – многоклеточный орган, имеющий одно- или многослойную стенку. Внутри многоклеточного спорангия формируется образовательная ткань – археспорий, из которого в результате мейоза (редукционного деления) образуются гаплоидные споры (спорогенез). Из спор при прорастании возникает гаплоидный организм – гаметофит, не идентичный диплоидному материнскому организму (спорофиту). У части высших растений (мхи, хвощи, некоторые плауны и папортники) все споры одинаковы по размерам и физиологическим особенностям. Это равноспоровые организмы. Из их спор возникают обоеполые гаметофиты. У других высших растений споры различаются по размерам и физиологическим особенностям (микроспоры и мегаспоры), - это разноспоровые организмы. Более мелкие и обычно более многочисленные микроспоры образуются в микроспорангиях, более крупные и малочисленные мегаспоры – в мегаспорангиях. Микроспоры, прорастая, дают начало однополому мужскому гаметофиту, на котором возникают мужские половые органы – антеридии. Мегаспоры при прорастании образуют женский гаметофит, несущий женские половые органы – архегонии. Разноспоровость встречается у некоторые плауновидных и папоротников, у всех голосеменных и покрытосеменных. Гаметофиту плауновидных, хвощевидных и папортниковидных представлен заростком - маленьким (от нескольких мм до 3 см), не расчлененным на органы растеньицем, живущим несколько недель (у плаунов – несколько лет) независимо от спорофита. На заростках в мужских половых органах (антеридиях) развиваются мужские половые клетки – сперматозоиды, которые, плавая в каплях воды, достигают женских половых органов (архегониев) и сливаются с яйцеклеткой. Гаметофиты могут быть однодомными, когда они формируют сразу и антеридий, и архегоний, либо однодомными (с половыми органами лишь одного типа). Благодаря крошечным размерам гаметофитов процесс оплодотворения у хвощей, плаунов и папоротников может происходить даже при ничтожно малых количествах воды в виде капель росы, тумана и др. Обычно при обоеполом гаметофите сперматозоиды и яйцеклетки созревают неодновременно, что снижает вероятность самооплодотворения.
13. Селагенелловые. Общая характеристика. Особенности строения спорофита в связи с условиями жизни. Строение гаметофитов. Биологическое значение разноспоровости. Селагенелловые. Многолетние травянистые многоосевые растения, удлиненные стебли несут чешуевидные листья. Латеральные меристемы (камбий, феллоген) отсутствуют. Порядок представлен в современной флоре полиморфным родом селагинелла или плаунок. Это наземные травянистые растения, реже лианы или эпифиты. Стебли не имеют признаков вторичного роста. В одних случаях растения не превышают 8-12 см, некоторые лианы до 18-20 м. Стебли прямостоячие или лежачие. От лежащих стеблей отходят вертикальные побеги. Стебель покрыт однослойной эпидермой. Устьица отсутствуют. Под эпидермой расположена гиподерма, выполняющая функции эпидермы и механическую функцию. Древесина из лестничных трахеид, луб из ситовидных трубок. Все корни придаточные. Гаметофиты раздельнополые, мелкие, симбиоза с грибами не образуют. Общее число видов 700. Распространены во влажных тропических лесах. По внешнему строению селагинеллы разнородны. На верхушке некоторых побегов развиваются спороносные колоски. Листья спороносного колоска – спорофиллы – заметно отличаются от вегетативных. В пазухах спорофиллов на короткой ножке развивается мега – и микроспорангии. Спорофилы несут язычки, которые никогда не отваливаются. Важное отличие селагенелловых от плауновых – их разноспоровость: мегаспорангии содержат 4 мегаспоры, микроспорангии – множество микроспор. Освободившись от спорангиев микро- и мегаспоры прорастают в благоприятных условиях. Мужской и женский гаметофиты развиваются в границах оболочек микро- и мегаспоры. Лишь небольшие лопасти заростка выдвигаются наружу. На лопастях женского заростка развиваются архегонии. Из оплодотворенной яйцеклетки развивается зародыш. При разноспоровости наиболее радикальная редукция гаметофита (заростка) – гаплоида и развитие до гигантских размеров спорофита – диплоида. Разноспоровость вызвала раздельнополость, т.е. двудомность заростков.
14. Порядок Хвощи. Особенности морфологии и анатомии. Признаки ксераморфной и гигроморфной организации у хвощей. Особенности строения эпидермы и устьиц. Стробилы. Цикл воспроизведения. Строение гаметофитов. Хвощевидные – это самый малочисленный отдел среди папоротникообразных. Единственный род Хвощ включает 32 вида, из которых 17 встречаются на территории нашей страны. Хвощи растут на болотах, лугах, в лесах и водоемах. Широко распространены по всему земному шару, за исключением Австралии и Новой Зеландии. Наибольшего развития хвощевидные достигали в каменноугольном периоде. Отмершие остатки древовидных хвощей того времени сформировали обширные запасы каменного угля. Современные представители отдела – многолетние травянистые корневищные растения. Их стебель высотой до нескольких десятков сантиметров расчленен на длинные междоузлия и узлы, что определило другое название этого отдела – Членистые. От узлов наземного побега отходят мутовки боковых побегов, которые часто неправильно принимают за листья. Настоящие листья хвощей мелкие чешуевидные, очень сильно редуцированы и не имеют хлорофилла. Они срастаются между собой, образуя трубку, которая охватывает междоузлие. Растущие боковые побеги прорывают эту трубку. Функцию фотосинтеза у хвощей берет на себя стебель. Его хлорофиллоносная ткань располагается в периферической части первичной коры. Клетки эпидермы и механической ткани хвощей способны накапливать кремнезем, что придает растениям повышенную прочность. В центральной части стебля молодых растений располагается сердцевина, которая постепенно разрушается. Поэтому в зрелых надземных побегах в центре находится полость, заполненная воздухом. По характеру надземных побегов, по их консистенции и функциональной морфологии виды хвоща можно разбить на 2 группы. У одних видов все надземные побеги однотипного строения. Они очень жесткие, обычно вечнозеленые и развивают верхушечные стробилы. У видов второй группы побеги двух типов – одни спороносные, буроватые или зеленые, а другие вегетативные, зеленые. Побеги обоих типов появляются весной, и они нежнее по своей консистенции и к зиме отмирают. На конце растущего вегетативного побега хвоща под защитой молодых листовых влагалищ расположена верхушечная клетка, имеющая вид трехгранной пирамиды, в результате деления на стебле возникают кольцевидные зачатки влагалищ, которые сохраняют способность к делению, образуя зону интеркалярной меристемы. В состав оболочки клеток эпидермы и в других тканях, но в меньшей степени, входит наряду с целлюлозой кремнезем. На поверхности стебля хвоща в области междоузлия видны ребрышки (гребни) и ложбинки. Число гребней связано с диаметром стебля.
Анатомическое строение хвощей различных видов довольно однотипно. Вдоль стеблей тянутся более или менее выраженные ребрышки и ложбинки. Под эпидермой в ребрышках, иногда в ложбинках, реже кольцом располагаются участки механической ткани. Между механической тканью находится паренхима первичной коры, содержащая хлоропласты и выполняющая функцию фотосинтеза. Во внутренней части коры проходят воздухоносные полости. Центральный осевой цилиндр нечетко отграничен от первичной коры, большая его часть приходится на сердцевину. В молодом стебле она состоит из живой паренхимы, которая быстро разрушается и образующаяся полость заполняется воздухом. Наличие большого количества межклетников свидетельствует, по-видимому, о том, что древние виды, как и многие современные виды хвощей, жили в болотистых местообитаниях. По периферии стелы тянутся закрытые коллатеральные пучки. Флоэма состоит из ситовидных трубок и паренхимы. В ксилеме вся протоксилема и большая часть метаксилемы разрушается. На их месте образуется узкая полость, по которой перемещается вода. Остатки метаксилемы обнаруживаются в виде кольчатых или спиральных трахеид. Пучки в междоузлиях идут параллельно друг другу. Входя в узел, сердцевина которой заполнена паренхимой, каждый пучок разветвляется на 3. Средний пучок входит в лист, а каждое боковое ответвление одного пучка сливается с боковым ответвлением соседнего пучка. Вновь сформированный пучок входит в следующее междоузлие. Таким образом, пучки одного междоузлия чередуются с пучками соседнего междоузлия. Устьица в междоузлиях приурочены к участкам эпидермы, которые выстилают ложбинки. Листовые влагалища, охватывающие основание каждого междоузлия, защищают зоны вставочной меристемы. По верхнему краю влагалища сидят зубцевидные листовые пластинки. Форма влагалища, цвет, форма и время жизни зубцов являются важным диагностическим признаком при определении видов.
Проводящая система стебля Хвощевидных представлена актиностелой или артростелой, то есть членистой стелой, состоящей из чередующихся между собой на протяжении стебля участков различного строения. Проводящие элементы ксилемы представлены трахеидами разного типа, а у хвощей также и сосудами; флоэма состоит из ситовидных элементов и паренхимных клеток. Характерной чертой Хвощевидных является наличие у них своеобразных несущих спорангии структур – спорангиофоров, отличных по строению от спорофиллов других растений. Мутовки спорангиофоров или образуют на стебле спороносные зоны, чередующиеся с обычными вегетативными листьями, или сидят на концах осей, образуя чистые (только из спорангиофоров) или смешанные (со стерильными листьями) стробилы. На подземном корневище образуются придаточные корни. Некоторые укороченные междоузлия подземного побега превращаются в клубеньки, в клетках которых в большом количестве откладывается крахмал. Современные хвощи не способны к вторичному утолщению стебля, поэтому древовидных форм в настоящее время среди них нет. Крупные хвощи, обитающие во влажных тропических лесах, хотя и достигают высоты до 10 м, вынуждены использовать в качестве опоры деревья. У большинства видов на вершине главного, а иногда и на боковых побегах формируются спороносные колоски стробилы, в которых развиваются спорангии. Споры образуются в результате редукционного деления, как правило, имеют шаровидную форму и содержат хлоропласты. Все современные хвощи – равноспоровые растения. Попав в благоприятные условия, спора прорастает в зеленый заросток (гаметофит) размером несколько миллиметров. У хвощей встречаются обоеполые, мужские и женские заростки. Формирование всех типов заростков из морфологически одинаковых спор является результатом так называемой физиологической гетероспории. Какой тип заростка образуется из споры, в основном определяется условиями внешней среды, в частности освещенностью. В антеридиях развиваются многожгутиковые сперматозоиды, в архегониях – яйцеклетки. Оплодотворение осуществляется при наличии капельно-жидкой воды (в дождливую погоду или во время обильной росы). Зигота начинает делиться без периода покоя, образуя зародыш, из которого вырастает новое бесполое поколение – спорофит. Размножаются хвощи и вегетативно – корневищами, образуя обширные заросли. Это многолетнее корневищное растение, встречающееся на полях и лугах. Ежегодно из почек,
15.Порядок сальвиниевые. Общая характеристика, жизненный цикл. Порядок Сальвиниевые характеризуется разноспоровостью и наличием шаровидных сорусов, открыто сидящих на подводных листьях. Оно включает один род сальвинию с 8 видами пресноводных растений. Из них 6 видов распространены в тропиках и субтропиках, 2 вида встречаются в умеренных областях; в России - в средней полосе, в Сибири, на Дальнем Востоке, на Кавказе и в Средней Азии. За время своего длительного существования в водной среде (сальвиниевые известны с мелового периода) сальвиния приобрела множество специализированных признаков. Все виды сальвиний - небольшие папоротники , размером до 15-20 см, живущие в стоячих либо медленно текущих водах. Тропические виды - многолетние растения, а виды умеренных областей - однолетники. Они имеют стелющиеся по поверхности воды побеги с мутовчатым листорасположением. Корни отсутствуют. В каждой мутовке из 3 листьев 2 листа плавающие, с цельной листовой пластинкой, а третий - подводный. Плавающие листья овальной или округлой формы; они покрыты восковым налетом и имеют на поверхности сосочки и волоски, что делает их несмачиваемыми. Подводный лист почти до основания рассечен на множество нитевидных долей, густо покрытых жесткими волосками. Выделяющиеся при дыхании пузырьки углекислого газа задерживаются между волосками, уменьшая удельный вес растения. Подводные листья своими нитевидными долями очень напоминают корни. На коротких черешках подводных листьев формируются шаровидные сорусы . Таким образом, подводные листья выполняют функции спороношения, всасывания воды и роль поплавка. Все части растения снабжены густой сетью воздухоносных полостей. В стебле расположена сифоностела с тонким цилиндром ксилемы; механические ткани очень слабо развиты. Виды сальвинии обладают способностью к вегетативному размножению за счет почек, которые могут формироваться между листьями. Развивающиеся из почек боковые побеги легко отламываются и дают начало новым растениям, в результате чего обширные водные территории могут быть сплошь покрыты сальвинией. Это часто мешает судоходству и рыбной ловле. У сальвинии плавающей (Salvinia natans) сорусы имеют шаровидную форму. В них на разросшейся плаценте располагаются либо микроспорангии , либо мегаспорангии . С поверхности сорусы покрыты двуслойным индузиумом , его слои соприкасаются только в основании и на самой вершине, а на остальном протяжении они разделены воздухоносной полостью, уменьшающей удельный вес сорусов.Спорангии имеют однослойную стенку, лишенную кольца. Выстилающий слой - тапетум , формирующийся из периферических археспориальных клеток, к моменту созревания спор расплывается и становится пенистым. После его затвердения образуется легкая пористая масса наподобие пенопласта, называемая массулой . В массулу погружены споры. В микроспорангиях образуются 64 микроспоры, а в мегаспорангиях полного развития достигает лишь 1 мегаспора. К осени растения отмирают, а тяжелые сорусы, богатые крахмалом, опускаются на дно и там зимуют. Весной стенки сорусов сгнивают, крахмал расходуется на прорастание спор, и легкие из-за массулы спорангии всплывают на поверхность воды. Микроспоры при прорастании развивают мужской гаметофит -заросток ( проталлиум ); он состоит из 2 вегетативных клеток , соответствующих вегетативному телу гаметофита. На нем образуются 2 антеридиальные клетки, каждая из которых формирует сильно редуцированный антеридий . Антеридий имеет стенку из 2 клеток и 1 сперматогенную клетку, дающую 4 сперматозоида. Мегаспора при прорастании образует менее редуцированный женский гаметофит, состоящий из множества вегетативных, т.е. проталлиальных, клеток. Под давлением заростка оболочка мегаспоры лопается и заросток выступает за ее пределы. Освободившаяся часть заростка зеленеет, и на ней формируется 3-5архегониев . После оплодотворения, которое происходит вслед за разрушением стенки мегаспорангия, из зиготы развивается зародыш. Благодаря гаустории он долго сохраняет связь с гаметофитом. Первый возникающий лист имеет щитковидную форму, следующие 2 листа округлые, густо покрытые волосками, и уже позднее формируется мутовка из 3 листьев. Семейство азолловые (Azollaceae) содержит 1 род азоллу (Azolla) с 6 видами, распространенными в тропиках и умеренных областях Северной Америки. Это маленькие нежные папоротники, похожие наюнгерманниевые печеночники . Сальвиниевые и азолловые обнаруживают много общего и в своем происхождении связаны, по-видимому, с древними вымершими гименофилловыми .
16. Стелярная теория. Понятие о стеле. Основные типы стел в микрофильной линии эволюции споровых растений. Уже в конце прошлого века в результате трудов фон Моля, де Бари и, главным образом, Сакса сложилось представление о том, что стебель растения состоит из трех систем тканей: покровные ткани, основная перенхима и погруженные в паренхиму проводящие пучки. Но с позиций такого членения трудно было описать и систематизировать разнообразные типы строения стебля, невозможно было сформулировать и четкую концепцию их эволюционного становления. Эти затруднения были преодолены стелярной теорией, сформулированной в основных чертах французским ботаником Ваном-Тигемом. Под стелой Ван-Тигель обозначил совокупность проводящих, арматурных и паренхимных тканей, ограниченных перициклом. Первоначально это чисто описательное понятие было введено для корня, впоследствии его распространили и на стебель. Изучение разных типов стелярной организации представляет большой интерес для понимания путей эволюции высших растений. Наиболее примитивным типом стелы является та, в которой проводящая ткань образует сплошную массу, и центральное ядро, состоящее из ксилемы, полностью окружено флоэмой. Такая стела называется протостелой. Протостела не только проста по структуре, но и представляет собой исходный тип, от которого в процессе эволюции произошли все другие типы стелы. Она была характерна для риниофитов и многих других вымерших форм. В современной флоре протостелу имеют некоторые плауны и папоротники. Наиболее примитивной формой протостелы является гаплостела (греч. гаплос - простой), состоящая из центрального пучка первичной ксилемы, окруженного цилиндром флоэмы. Более продвинутой формой является актиностела (греч. актинос - луч), имеющая радиальные выступы ксилемы в виде В процессе эволюции протостела дала начало сифоностеле (от греч. сифон ≈ трубка). Сифоностела имеет трубчатое строение, в ней появляется сердцевина. Возникновение сифоностелы сделало возможным существование более крупных организмов. Ксилема, играющая еще и роль арматурной ткани, перемещается на периферию стебля, образование такой трубчатой конструкции делает стебель еще более прочным. Различные типы сифоностелы характерны для многих папоротников. Дальнейшее усложнение стелы связано с появлением у папоротников крупных листьев (макрофильная линия эволюции). Сифоностела как бы дробится на отдельные участки. В результате дробления возникает диктиостела(греч. диктион - сеть) папоротников и эустела (настоящая стела) семенных растений. Последнее звено в эволюции стелы стебля представляет атактостела (греч. а ≈ отрицание, тактос ≈ порядок). Она характерна для однодольных растений. От эустелы отличается отсутствием в проводящих пучках камбия. Сами же проводящие пучки как бы беспорядочно разбросаны по всему поперечному сечению среза. Выявляя существенные различия между основными отделами высших растений по архитектонике их проводящей системы, Стелярная теория имеет большое значение не только для анатомии растений, но и для познания филогенеза растений в целом
17. Порядок Многоножковые. Общая характеристика. Морфологические разнообразия. Жизненные формы. Варианта строения и расположения сорусов и спорангиев. Жц. 18. Общая характеристика и классификация семенных растений. Особенности морфологического и анатомического строения семенных растений. Семенные растения по сравнению с споровыми представляют собой более высокий уровень организации, поскольку главным зачатком для расселения вида является качественно новое образование – семена. Эволюционное преимущество семенных растений перед споровыми заключается в том, что половой процесс у них не зависит от капельно-жидкой воды. Благодаря такой независимости семенах растения получили возможность расселяться на всей Земле и стали прогрессивной группой. В отличие от высших споровых, в которых споры одноклеточные, в семенных растений семена многоклеточное и содержит в себе сформирован зародыш и запас питательных веществ для его развития. В семенных растений наблюдается дальнейшее усовершенствование и еще большее преимущество в цикле развития спорофита и редукция гаметофита, существование которого полностью развития спорофита и редукция гаметофита. Семенные растения, появившиеся около 360 млн лет назад, – господствующая в настоящее время группа высших растений. Семенные растения подразделяют на 2 отдела: голосеменные размножающиеся семенами, но не образующие плодов, и покрытосеменные -имеющие семена, заключенные в плоды. Эволюция шла в направлении редукции гаметофита и дальнейшего развития спорофита. Семя - новый орган, возникший в процессе эволюции для размножения растений. Семя содержит зародыш будущего растения – спорофита. В состав зародыша входят зародышевый корешок, стебелек, почечка и зародышевые листья (семядоли). Зародыш защищен семенной кожурой и имеет запас питательных веществ; в отличие от этого в одноклеточной споре запас питательных веществ небольшой и она быстро теряет всхожесть. Семя имеет приспособления для расселения, проходит период покоя при прорастании, т.е. процесс прорастания наступает при более благоприятных условиях. Спорофит получил полное господство в цикле развития над гаметофитом. Гаметофит крайне редуцирован, утратил самостоятельное существование и полностью зависит от спорофита, на котором он образуется и формируется. Половой процесс у семенных растений не связан с капельножидкой средой. Благодаря такой независимости они получили воз- можность расселиться по всей поверхности Земли и стать господствующей группой растений. Важным прогрессивным приспособлением, исключающим зависимость процесса оплодотворения от водной среды, явилось воз- никновение в процессе эволюции пыльцевой трубки, доставляющей мужские гаметы к яйцеклетке.
19. Голосеменные растения. Морфолого-анатомическое строение. Принципиальный цикл воспроизведения. Строение стробилов, мужского и женского гаметофита на примере сосны. Голосеменные – древняя группа растений, появившаяся еще в девоне. Голосеменные представляют собой растения с более высоким уровнем организации, чем высшие споровые, так как средством для их расселения является семя. Характерные особенности голосеменных: 1) разноспоровые растения; 
12 Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|


 На верхушке некоторых побегов развиваются спороносные колоски. Листья спороносного колоска – спорофиллы – заметно отличаются от вегетативных. В пазухах спорофиллов на короткой ножке развивается мега – и микроспорангии. Спорофилы несут язычки, которые никогда не отваливаются. Важное отличие селагенелловых от плауновых – их разноспоровость: мегаспорангии содержат 4 мегаспоры, микроспорангии – множество микроспор. Освободившись от спорангиев микро- и мегаспоры прорастают в благоприятных условиях. Мужской и женский гаметофиты развиваются в границах оболочек микро- и мегаспоры. Лишь небольшие лопасти заростка выдвигаются наружу. На лопастях женского заростка развиваются архегонии. Из оплодотворенной яйцеклетки развивается зародыш. При разноспоровости наиболее радикальная редукция гаметофита (заростка) – гаплоида и развитие до гигантских размеров спорофита – диплоида. Разноспоровость вызвала раздельнополость, т.е. двудомность заростков.
На верхушке некоторых побегов развиваются спороносные колоски. Листья спороносного колоска – спорофиллы – заметно отличаются от вегетативных. В пазухах спорофиллов на короткой ножке развивается мега – и микроспорангии. Спорофилы несут язычки, которые никогда не отваливаются. Важное отличие селагенелловых от плауновых – их разноспоровость: мегаспорангии содержат 4 мегаспоры, микроспорангии – множество микроспор. Освободившись от спорангиев микро- и мегаспоры прорастают в благоприятных условиях. Мужской и женский гаметофиты развиваются в границах оболочек микро- и мегаспоры. Лишь небольшие лопасти заростка выдвигаются наружу. На лопастях женского заростка развиваются архегонии. Из оплодотворенной яйцеклетки развивается зародыш. При разноспоровости наиболее радикальная редукция гаметофита (заростка) – гаплоида и развитие до гигантских размеров спорофита – диплоида. Разноспоровость вызвала раздельнополость, т.е. двудомность заростков. лучей (на поперечном срезе имеет вид звезды). При переходе к актиностеле флоэма и ксилема имеют большую поверхность соприкосновения с окружающими живыми тканями, что способствует лучшему проведению веществ.
лучей (на поперечном срезе имеет вид звезды). При переходе к актиностеле флоэма и ксилема имеют большую поверхность соприкосновения с окружающими живыми тканями, что способствует лучшему проведению веществ.