|
|
Альтернативные формы двойной спирали ДНКВсе, о чем мы говорили, касалось наиболее распространенной, так называемой В-формы двойной спирали ДНК. Известны также два других изомерных типа двойной спирали. Они образуются благодаря тому, что валентные углы между основаниями и сахаром могут меняться, а дезоксирибозное кольцо и сахарофосфатный остов достаточно гибки, чтобы могли сформироваться альтернативные конфигурации. Редко встречающаяся А-форма, существующая только при пониженной влажности, отличается от В-формы тем, что плоскости оснований составляют с перпендикуляром к оси спирали угол 20° :
Поэтому расстояние между парами оснований по вертикали уменьшается до 0,29 нм, а число пар на виток увеличивается до 11—12. Какова биологическая функция А-формы ДНК—пока неясно. Характерной особенностью В-формы ДНК является то, что сахарофосфатные остовы обеих цепей образуют правую спираль. Однако при определенных условиях участки ДНК, для которых характерно чередование пуриновых и пиримидиновых нуклеотидов, принимают форму левой спирали. При этом расстояние между соседними парами оснований увеличивается до 0,77 нм, а число пар на один виток—до 12. Остов молекулы ДНК имеет зигзагообразный вид, поэтому подобная форма получила название Z-ДНК. Вопрос о том, существует ли Z-ДНК в естественных условиях и образуется ли она в определенных участках В-спирали под действием специфических белков, способных переводить В-форму в Z-форму, сейчас интенсивно исследуется.
Размер молекул ДНК Обычно размер молекулы ДНК выражается в числе пар нуклеотидов, при этом за единицу берется тысяча пар нуклеотидов (т.п.н.). Мол. масса одной т.п.н. В-ДНК равна в среднем 6,6*105, а ее длина составляет 340 нм. Если принять все необходимые меры, чтобы не разрушить ДНК при выделении, и использовать мягкие методы измерения длины, то обнаружится удивительное соответствие между длиной молекулы ДНК и массой одной небольшой хромосомы. Разнообразие форм ДНК Существовавшее до недавнего времени мнение о том, что В-ДНК — это совершенная двойная спираль, геометрия которой одинакова независимо от нуклеотидной последовательности, в действительности не совсем корректно. Детальный рентгеноструктурный анализ, построение моделей и термодинамические расчеты показали, что плоскости соседних пар оснований не строго параллельны. Каждая комплементарная пара оснований является как бы клином, отклоняющим ось спирали в одном или в другом направлении. Наибольший «крен» наблюдается тогда, когда два соседних аденина в одной цепи спарены с двумя тиминами другой. В этом месте происходит локальное искривление спирали. Если такие пары встречаются с периодичностью примерно один раз на 10 пар (т.е. один раз на каждый виток спирали), то молекула ДНК приобретает заметно искривленную форму. Изгибы в молекуле ДНК наблюдаются в тех участках последовательности, где с необычно высокой частотой встречаются повторы (А•Т)5–6, разделенные GC-богатыми участками из четырех-шести нуклеотидов. Биологическая роль искривления ДНК окончательно не установлена. Предрасположенность к такому изгибанию, зависящая от последовательности оснований, может иметь значение при наматывании молекулы ДНК на гистоновые октамеры в хроматине. Возможно, изгибание ДНК существенно и при специфическом связывании ДНК с белками в процессе регуляции экспрессии генов.
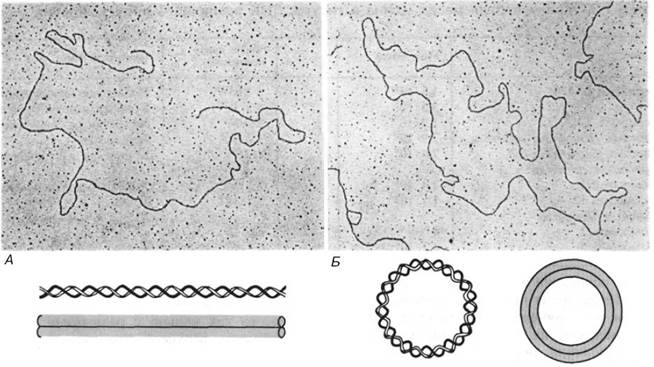
ДНК может находиться в линейной или кольцевой форме:
Бактериальные плазмиды, хромосомы некоторых бактерий, большинство митохондриальных и хлоропластных ДНК, геномы вирусов млекопитающих представлены единственной ковалентно замкнутой кольцевой дуплексной молекулой ДНК. Хромосома бактериофага λ на разных стадиях жизненного цикла существует то как линейная молекула, то как замкнутая кольцевая структура, то как кольцо с разрывами. По-видимому, никакого верхнего предела для размера кольцевой двухцепочечной молекулы ДНК не существует. ДНК в клетке обычно находится в комплексе с белками. Связанный белок слегка раскручивает спираль ДНК, соответственно и число витков спирали на единицу длины становится меньше, чем у свободной В-ДНК. При удалении белка восстанавливается обычное число правозакрученных (положительных) витков спирали. В линейной молекуле ДНК это происходит достаточно легко, поскольку обе цепи свободно вращаются одна вокруг другой. В замкнутой же кольцевой молекуле общее число витков спирали топологически фиксировано, и число оборотов одной цепи вокруг другой не может быть изменено без компенсаторного образования витков противоположного знака где-нибудь в другом месте молекулы. Итак, когда естественные кольцевые дуплексы освобождаются от белков, с которыми они часто бывают связаны in vivo, происходит следующее: 1) число правозакрученных (положительных) витков спрали возрастает до величины, характерной для В-ДНК; О таких молекулах говорят, что они обладают отрицательной сверхспиральностью:

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|