|
|
ФУНКЦИИ МОЗГА: МЕХАНИЗМ ДВУХ ПРОЦЕССОВKARL H.PRIBRAM Stanford university LANGUAGES OF THE BRAIN Experimental paradoxes And principles In neuropsychology Prentice-hall, inc. Englewood cliffs, new jersey. К.ПРИБРАМ ЯЗЫКИ МОЗГА Экспериментальные Парадоксы и принципы Нейропсихологии Перевод с английского Я. Н. Даниловой и Е. Д. Хомской Под редакцией и с предисловием действительного члена АПН СССР А. Р. Лурия Издательство «Прогресс» Москва 1975 Редакция литературы по философии Переводна русский язык с изменениями. «Прогре И10508'796 9-74 006(01)75 ПРЕДИСЛОВИЕ РЕДАКТОРА РУССКОГО ИЗДАНИЯ Предлагаемая советскому читателю книга принадлежит перу одного из наиболее творческих представителей американской нейропсихологии — профессору Стэнфордского университета К. Прибраму. Автор этой книги начал свою деятельность нейрохирургом и затем перешел к изучению функций мозга животных. Вместе со своими многочисленными сотрудниками он провел большое число исследований, в которых он пытался выяснить, какую роль в поведении животного играют отдельные структуры головного мозга и как изменяется поведение животного после их разрушения. Работы автора, посвященные функции лобных долей и выработке и удержании программ, направляющих поведение животного, и роли задних (височно-затылочных) отделов мозга в переработке поступающей информации, хорошо известны психологам и физиологам; именно эти работы и выдвинули К. Прибрама на место одного из ведущих авторитетов нейропсихологической науки. К. Прибрам известен советскому читателю и по ряду его выступлений, получивших широкий отклик. Написанная им совместно с Дж. Миллером и Ю. Галантером книга «Планы и структура поведения» была издана в Советском Союзе (изд-во «Прогресс», М., 1965). В ней был выдвинут новый подход к анализу поведения животных, исходящий из учета той роли, которую играют в поведении «Образы», создаваемые при переработке получаемой информации, и «Планы», лежащие в основе активной деятельности животных и человека. Концепции, предложенные авторами этой книги, близкие к современной кибернетике и учению о саморегулирующихся функциональных системах, развитому в нашей стране П. К. Анохиным, получили большое распространение и были положительно оценены советской научной общественностью. Признание работ К. Прибрама привело к тому, что ему было поручено наряду с виднейшими учеными-психологами выступить с обобщением итогов всех психофизиологических докладов, которые были представлены на XVIII Международном психологическом конгрессе в Москве. Книга К. Прибрама «Языки мозга» носит совсем особый характер. С одной стороны, она отличается очень большой широтой и новизной подходов к проблемам мозговой организации психических процессов. Автор привлекает к разрабатываемой им концепции мозговой деятельности новейшие данные физики (голографии), молекулярной биологии, морфофизиологии нейронных структур, учение о саморегулирующихся системах и кибернетику. В этом отношении книга лишена всякой традиционности и не столько резюмирует уже накопленные в науке данные, сколько пытается проложить новые пути, которые, по всей вероятности, будут с достаточной полнотой разработаны лишь будущими поколениями исследователей; однако уже сейчас тот интересный новый материал, который предлагает автор, и уникальные по своей выразительности иллюстрации, которые читатель найдет с первых страниц книги, придают ей большую свежесть и дают важнейшую информацию о последних достижениях в этой области науки. Пусть многие из высказываемых автором гипотез отличаются чрезмерной смелостью и иногда являются спорными; книга толкает на новые поиски, заставляет пересмотреть многие уже устоявшиеся концепции, и в этом — будящем мысль воздействии — несомненно, одно из основных достоинств книги. С другой стороны, эта книга, как и другие публикации автора, представляет бесспорный интерес в том отношении, что она отражает попытки выйти из того глубокого кризиса, в котором оказалась американская научная мысль, в течение десятилетий испытывавшая тормозящее влияние упрощенных и механистических схем бихевиоризма. Как и в прошлой книге, написанной совместно с Дж. Миллером и Ю. Галантером, автор решительно отходит от бихевиористского понимания поведения как цепи стимулов и реакций; он столь же решительно отбрасывает самую мысль о возможности игнорировать проблемы сознания, с одной стороны, и анализ физиологических механизмов, лежащих в основе сознательной деятельности, — с другой. Пусть самая возможность понять сознательную деятельность как продукт сложнейшего общественно-исторического развития еще чужда автору (который в большой мере остается биологом и пытается биологически и нейрофизиологически трактовать механизмы, лежащие в основе психической деятельности). Пусть он иногда определяет свою позицию как позицию «субъективного бихевиоризма» (иначе говоря, изучение поведения, не исключающее, а предполагающее анализ субъективного мира и ищущее. «го нейрофизиологических объяснений); пусть он высказывает ряд положений, которые, очевидно, вызовут острые дискуссии. Однако его решительный отход от упрощенных, механистических схем бихевиоризма, попытки проложить новые пути в нейропсихологии представляют несомненный интерес, и именно эти попытки дают читателю возможность увидеть контуры будущей психофизиологии. Остается пожалеть, что автор недостаточно знаком с успехами советской физиологии (в частности — с работами П. К. Анохина, ряд положений которого почти полностью совпадает с его взглядами) и с теорией деятельности, развиваемой в советской психологической науке. Более близкое знакомство с данными обеих дисциплин существенно помогло бы ему уточнить ряд высказываемых им идей. Можно не сомневаться в том, что книга К. Прибрама, в которой автор пытается разобраться в том, на каких «языках» говорит мозг и в каких параметрах выражается его деятельность, будет с пользой прочитана советскими исследователями и послужит для них толчком к новым продуктивным размышлениям. А. Р. Лурия © «Прогресс», 1974
ПРЕДИСЛОВИЕ АВТОРА К АНГЛИЙСКОМУ ИЗДАНИЮ Появление книги «Языки мозга» вызвано несколькими обстоятельствами. Во-первых, существует профессиональная потребность сформулировать для самого себя и своих коллег научные взгляды, которые сложились во время моей работы и сейчас направляют ее. Таким образом, являясь прежде всего теоретическим исследованием, книга содержит анализ тех парадоксов и загадок, которые неожиданно возникали в ходе экспериментов. В то время, когда проводилось исследование, ряд фактов не поддавался объяснению в рамках теории, разделяемой большинством исследователей. Поскольку попытки найти объяснение таким парадоксам придавали своеобразие научным достижениям лаборатории, изучающей отношение между мозгом и поведением, я постарался сохранить его в этой книге. Но не одни лишь профессиональные интересы побудили меня написать эту книгу в ее настоящем виде. Обилие новых данных и важность разработки проблем взаимоотношения мозга, поведения и сознания вызвали в последние годы интерес к этим проблемам со стороны широкого круга людей. К ним относятся не только специалисты по психологии или неврологии, но и молодые, начинающие исследователи, студенты, инженеры и физики, биологи и биохимики, социологи и психиатры, философы и др. Такая аудитория требует чего-то большего, чем «самые последние данные о базолатеральном ядре амигдалы» или «различии между задачами на активное и пассивное избегание». Интуитивно она догадывается, что открытия в области изучения мозга м поведения важны не только «для науки», но и для самих читателей. Эта книга и является попыткой разрешить многие «опросы, возникающие перед теми, кто, как и я, старается дать объяснение тем парадоксальным результатам, которые были получены при изучении мозга и поведения. Поэтому я попытался найти в этой книге такую манеру изложения, которая сочетала бы в себе описание деталей» интересующих профессионалов, с тем, что представляет всеобщий" интерес. Основное направление моих лабораторных исследований создает идеальные условия для выполнения этой задачи: цель моих экспериментов состоит в сущности в определении посредством поведенческого анализа функций различных систем нейронных структур, составляющих мозг. 9ia «системная» нейропсихология является связующим звеном между нейрофизиологией — электрическим и химическим изучением функций нервных клеток (и их частей)—и экспериментальной психологией — поведенческим анализом функций организма как целого. За последние годы появилась и третья линия исследований, связанная с использованием компьютеров при изучении многих интересующих нас проблем. Компьютеры помогают контролировать эксперименты, анализировать полученные данные и указывают новый путь для будущих исследований. Можно существенно сэкономить усилия исследователей, если дать какому-либо научному подходу «зеленую улицу», чтобы проверить пригодность тех или иных методов и подробно описать последствия их применения, которые далеко не очевидны, хотя и подразумеваются. Закон, выведенный с помощью компьютера, имеет для биолога, изучающего поведение, такое же значение, как эксперимент in vitro (в стеклянной пробирке) для биохимика. В обоих случаях такое моделирование способствует разработке системы научных понятий, с помощью которых можно адекватно анализировать процессы, протекающие в живых организмах. Поэтому такой «системный» подход к проблемам нейропсихологии волей-неволей предполагает знакомство исследователя с достижениями других дисциплин, имеющих свой собственный круг проблем и разработавших соответствующие понятия и свой подход к решению этих проблем. Короче говоря, нейропсихолог, стоящий на такой позиции, должен пользоваться разными «языками». Результаты моих собственных исследований подтвердили необходимость прибегать к множеству различных «языков». Очевидно, мозг организует перцептивные, моторные и мнестические процессы путем многократного перекодирования получаемой информации и многократной перестройки своей собственной активности. Сенсорные возбуждения трансформируются в динамические структуры нервной активности, которые позволяют сохранять информацию без ее чрезмерной потери. Дальнейшее преобразование в другие нервные структуры, в другие нервные «коды» происходит при последующей «переработке информации» и формировании сложных форм поведения. Поэтому многое в моей работе связано с анализом набора тех мозговых кодов, тех «языков» мозга, которые используются на той или иной фазе психологической переработки. Какие мозговые коды делают возможным опознание зрительного образа? Какие мозговые коды координируют постройку гнезда или искусное исполнение фортепьянной сонаты? Какие мозговые коды создают ощущение голода, сонливость, апатию или заинтересованность? Каковы те операции мозгового кодирования, которые позволяют мозгу одного человека общаться с мозгом другого? Что же представляют собой «языки мозга»? В этой книге предпринята попытка определить общие принципы мозговых кодирующих механизмов и преобразований, используемых при перекодировании. В ходе работы мы столкнулись с целым рядом парадоксов: экспериментальные результаты порой настолько отличались от того, что мы ожидали, что мы неизбежно должны были усомниться в правильности наших прежних представлений о работе мозга. Естественно, что это заставляло нас разрабатывать новые теории и предлагать новые гипотезы. Книга «Языки мозга» использует биологический язык химии, физиологии, неврологии, язык психологов, изучающих поведение, и язык инженеров, имеющих дело с компьютерами. Но поскольку эти языки нужно было сделать понятными для широкой аудитории, которая включает в себя как представителей различных дисциплин, так и людей, не являющихся специалистами, мы сохраняем в этой книге лишь основные понятия и слова всех этих языков. Книга состоит из четырех частей, каждая из которых относительно независима от других и может служить введением к последующей, — учет специфических интересов отдельных групп читателей мог бы продиктовать другую структуру книги. Итак, в книге «Языки мозга» излагаются принципы мозгового кодирования. В каждой из ее четырех частей исследуются различные проблемы. Первая часть посвящена основной функции мозга и «логике» построения нейронных структур, которые позволяют мозгу формировать применяемые им коды. Во второй части показана роль мозга в организации психических процессов. В ней анализируется процесс кодирования, участвующий в восприятии, мотивации и эмоции. В третьей части рассматриваются проблемы нейронного контроля и пластичности поведения. И, наконец, последняя часть посвящена структуре коммуникативных процессов в терминах знаков, символов и внутренней речи, которые регулируют действия человека. Таким образом, в книге показана необходимость мозга формировать различные коды, что приводит к появлению разных языков и превращает интеллектуальное сообщество в довременную Вавилонскую башню; вместе с тем она предоставляет и средства, позволяющие преодолеть это затруднение. В дискуссиях о тех качествах, которые делают человека человеком, исследователи часто пользуются дихотомиями: сознание — тело, сознание — машина, сознание — мозг, сознание — поведение. На страницах этой книги я попытался подойти к анализу таких дихотомий, исходя из того, что они являются выражением различных «языков мозга»_ Я благодарен прежде всего тем авторам, которые за последнее время написали ряд работ по физиологической психологии. Их успех помог мне найти свою собственную манеру изложения «Языков мозга». Это сделало книгу дополнением к общепринятому пониманию отношений между мозгом и поведением. В создании этой книги принимали участие многие исследователи. В ней цитируются работы тех, кто в течение ряда лег вместе со мной создавал лабораторию. Многие другие помогли» мне уточнить мои концепции в дискуссиях, подробные ссылки на их работы будут даны в следующей книге, за которую я теперь принимаюсь; там же будут освещены более специальные вопросы, разрабатываемые в моей лаборатории. «Известно, что когда Клерк Максвелл был: ребенком, у него была мания требовать, чтобы ему все объясняли; и, когда люди отделывались от него с помощью туманных словесных объяснений, он нетерпеливо прерывал их, говоря: «Да, но я хочу, чтобы вы мне сказали, что же из этого следует!» Поскольку его интересовала истина, то только прагматик мог бы сказать ему, что именно следует из сказанного... Истина возникает из фактов, но она и опережает факты, и кое-что к ним прибавляет, а эти факты вновь создают или открывают истину... и так до бесконечности. Между тем сами по себе «факты» не составляют истины. Они просто существуют. Истина состоит в той уверенности, которая начинается с фактов и кончается ими». Уильям Джеме
ЧАСТ1, ПЕРВАЯ ФУНКЦИИ МОЗГА: МЕХАНИЗМ ДВУХ ПРОЦЕССОВ «Язык, с помощью которого передается информация [в мозге] ... не соответствует и не должен соответствовать тому языку, которым люди пользуются в общении друг с другом». Питтс и Мак-Наллок
Глава I СОСТОЯНИЯ И ОПЕРАТОРЫ ВВЕДЕНИЕ «Я вас люблю». В Париже была весна, и эти слова были произнесены с прелестным скандинавским акцентом. Это событие произошло на заседании ЮНЕСКО, посвященном проблемам изучения мозга и человеческого поведения. Слова эти были произнесены не белокурой красавицей, а маленьким блестящим металлическим устройством, которое держал в руках известный психолингвист. Устройство поразило всех нас простотой своей конструкции. Громкоговоритель регулировался с помощью только двух ручек. Одна изменяла состояние электронной цепи, которое моделировало напряжение голосовых связок; другая регулировала импульсы, генерируемые цепью и имитировавшие взрывные звуки воздушной струи, ударявшейся по связкам. Может ли это простое устройство иметь отношение к изучению человеком самого себя? Нельзя ли вызывать и контролировать все поведение с помощью столь же простого нейронного механизма? Не является ли нервнйя система «двухкнопочным» механизмом двойного процесса, в котором один процесс выражается в терминах нейроэлектрических состояний, а другой — в терминах особых пульсирующих операторов, воздействующих на эти состояния? То, что нервная система фактически действует посредством импульсов, хорошо известно. Существование нейроэлектрических состояний мозга также было установлено, но доказательства их существования были получены не сразу, и понимание их значения для изучения поведения приходило лишь постепенно даже в нейрофизиологии. Поэтому в первой главе рассматриваются данные, свидетельствующие о правдоподобии двухпроцессной модели функций мозга. . Чтобы понять, в чем заключаются функции мозга, мы должны вначале понять, из каких единиц состоит нервная система. Классический анализ определяет эти единицы как нейроны — клетки, которые структурно и функционально изолированы друг от друга* мембранными оболочками. Однако такое упрощенное представление об изолированном нейроне как единственном организаторе мозговых процессов вызывает затруднение при попытке охарактеризовать нейроэлектрические состояния как часть такого двойного механизма. В двух первых разделах дается беглый обзор основ нейронной теории и тех причин, по которым в настоящее время в нее должны быть внесены некоторые поправки. В конце XIX в. в нейробиологии возникла полемика о том, состоит ли мозговая ткань из единиц — клеток, подобно всем другим тканям тела. Результаты этой полемики были настолько убедительны, что нейрон и его способность действовать как единица, разряжающаяся электрическим потенциалом по закону «все или ничего», перестали быть предметом теоретического рассмотрения. Еще никто не «видел» нейрон мозговой ткани; никто не проследил полностью его протяженность и не показал, что он действительно изолирован от других нейронов (рис. 1-1). Каким же образом нейронная теория получила в дальнейшем всеобщее признание и почему мы вновь вернулись к этой проблеме сегодня? Спор вокруг нейронной теории достиг своей кульминации в конце XIX в. Непрерывность нервной ткани признавали такие известные нейроанатомы, как Билыповский и Гольджи; Валытейер и Рамон-и-Кахал рассматривали нейрон как независимый, но соприкасающийся с другими нейронами. Однако убедительный анализ был дан не этими нейроанатомами, а сэром Чарлзом Шеррингтоном в его лекциях в Йельском университете, опубликованных как классическая работа «Интегративная деятельность нервной системы». Ранее Шеррингтон совместно с Фостером предложил термин «синапсы» для обозначения предполагаемых связей между отдельными нервными клетками. Он внес большой теоретический вклад в нейронную теорию, противопоставив нейрофизиологические данные, основанные главным образом на изучении электрических явлений в нервных стволах, в&щопсихологичеспим *, базирующимся на изучении рефлекторной деятельности у «спинального животного» (нашедшей отражение в концепции рефлекторной дуги, см. гл. V). Шеррингтон высказал мысль, что парадоксальные расхождения результатов нейрофизиологических и поведенческих исследований могут получить объяснение, если обратиться к нейроанатомическому описанию синапсов. Основные расхождения состояли в том, что (1) в изолированном стволе сигнал, вызванный стимуляцией какой-нибудь его точки, передается в двух направлениях, тогда как проведение сигнала при осуществлении спинального рефлекса происходит в одном направ- * Автор употребляет термин «neurobehavioral», который далее будет переводиться уже прочно вошедшим в литературу термином «нейропсихологический». — Прим. ред. Рис. 1-1. Сканирующая электронная микрофотография, показывающая расположение нервных волокон в сетчатке Necturus'a. Волокна (дендриты) берут начало от внутреннего членика колбочки и распространяются в сторону наружного. Следует отметить, что точки контакта необязательно находятся на нервных окончаниях (Lewis, 1970).
лении, и (2) скорость проведения импульса по нервному стволу значительно выше, чем скорость ответа, получаемого в результате рефлекса. Помимо этих, было отмечено девять других расхождений. Большинство из них касалось степени соответствия между адресованным организму стимулом и полученным ответом: соответствие всегда было большим для нервного, чем для поведенческого ответа. Чтобы объяснить эти расхождения, синапсам приписывались такие предполагаемые функции, как пространственная конвергенция и временная суммация, утомляемость, облегчение и торможение. Следующее утверждение Шеррингтона дает представление о его концепции: «Наиболее характерными особенностями проведения в рефлекторных дугах по сравнению с нервными стволами являются: 1) меньшая скорость, если судить по измерениям латентного периода между нанесением раздражения и появлением конечного эффекта, причем это различие больше для слабых раздражителей, чем для сильных; 2) менее близкое соответствие между моментом прекращения раздражения и моментом прекращения конечного эффекта (т. е. имеет место отчетливое «последействие»); 3) менее близкое соответствие между ритмом раздражения и ритмом конечного эффекта; 4) менее близкое соответствие между градуальным увеличением интенсивности раздражения и градуальным увеличением интенсивности конечного эффекта; 5) значительное сопротивление к прохождению нервного импульса, которое, однако, без труда преодолевается последовательностью импульсов (временная суммация); 6) необратимость направления проведения по сравнению с обратимостью в нервных стволах; 7) утомляемость в противоположность сравнительной неутомляемости нервных стволов; 8) более значительная изменчивость пороговых величин раздражения по сравнению с нервными стволами; 9) рефрактерный период,» проторение», торможение и способность впадать в состояние шока в степени, неизвестной для нервных стволов; 10) большая зависимость от кровообращения и снабжения кислородом (Ферворн, Винтерштейн, Бейер и др.); 11) более выраженная восприимчивость к действию различных лекарств-анестетиков» (Шеррингтон, 1969, стр. 40—41). Йороче говоря, результаты электрофизиологического исследования нервных стволов и поведенческого изучения рефлекса не совпадали. Нервы казались простыми проводящими путями для импульсов; рефлекс же обнаружил такую сложность организации, которая не могла быть объяснена простой «проволочной» моделью. Поэтому Шеррингтон принял «нейронную теорию», высказав предположение, что «провода», образующие нервную систему, составляют не непрерывную сеть, а слегка отделены друг от друга и включают структуру, которую он назвал синапсом. Затем он наделил синапс всей той сложностью, которую необходимо принимать во внимание при объяснении рефлекторного поведения. Таким образом, свойства изучаемого поведения были отнесены за счет свойств соединений между нейронами, а не их проводниковой части (рис. 1-2 и 1-3). Теоретическая гипотеза Шеррингтона в значительной степени игнорировалась исследователями в области нейрофизиологии и нейропсихологии. Внимание было сосредоточено на существова- нии и свойствах синапсов. Эти свойства детально изучались с помощью электронной микроскопии и записи электрической и Рис. 1-2. Скопление нейронов в сером веществе спинного мозга обезьяны, иллюстрирующее обычное расположение тел нервных клеток и их аксонных и дендритных отростков. Большая часть пространства между телами клеток занята нервным сплетением, состоящим из этих отростков и множества входящих и разветвляющихся аксонных окончаний. Сплетение аксонов и дендритов, посредством которого главным образом осуществляется связь одного нейрона с другим, известно под названием «нейропиль». р — перикарион; а — аксон; d — дендрит; пр — нейропиль; л — ядро нейрона большого размера, вероятно мотонейрона. Парафиновый срез, 1,5 |х, окраска по Бодиану (Х150). (Видимое пространство вокруг тел нервных клеток — артефакт в результате сморщивания ткани). (Bodian, 1967.) химической активности соединений. Однако интерпретация этих результатов почти не выходила за рамки вопроса: «Как осуществляется прохождение нервных импульсов через барьер синапса?» Ссылка на разряды, генерируемые клеткой, и на проводящие-
•двойства синаптических путей слишком часто рассматривалась как самоочевидная и достаточная. Немногие ученые придерживались тезиса Шеррингтона о том, что сложность поведения (и психологических процессов) следует объяснять сложностью организации соединительных (синаптических) механизмов в центральной нервной системе.
Рис. 1-3. Схема микроструктуры синаптических связей в коре. Частично перекрывающие друг друга окружности изображают область соединительных контактов между разветвлениями входных аксонов и кортикальных дендритов (Scheibel and Scheibel, см. Chow and Leiman, 1970). Цель данной книги состоит в том, чтобы восполнить этот пробел. В ней сделана попытка, исходя из идей Шеррингтона, решить проблему взаимоотношений между мозгом, сознанием и поведением, подобно тому как Шеррингтон решал проблему отношений между спинным мозгом и рефлексом. Проблемы, возникающие при изучении головного мозга, тоньше и сложнее, чем проблемы, связанные со спинным мозгом. Тем не менее можно уже теперь достигнуть значительных успехов в понимании этих проблем при осторожном использовании классического нейробиологического подхода Шеррингтона. Однако неврологи, физиологи и психологи часто его игнорируют, переходя от отчаяния к неопределенным надеждам, а порой даже вообще отрицают существование такой проблемы. Изучение отношений между мозгом, сознанием и поведение» может быть продуктивным только в том случае, если ставятся ограниченные проблемы. Каковы же эти проблемы? НЕКОТОРЫЕ НЕЙРОПСИХОЛОГИЧЕСКИЕ ФАКТЫ Хотя нейрофизиологический эксперимент основывался на нейронной теории, некоторые известные психологи неоднократно отмечали, что понимание нервной системы как функционирующей исключительно в виде набора дискретно проводящих единиц не согласуется с полученными ими экспериментальными данными. Эти исследователи объясняли свои наблюдения, прибегая скорее к той или иной модели поля, чем к квантовой, дискретной, вероятностной нейронной теории. Разногласия, коротко говоря, сводились к следующему: «Физиолог слепо верит в то, что, поскольку мозг состоит из нейронов, он способен только к такому возбуждению, которое является суммой возбуждений многих нейронов, и что эти центральные нейроны подчиняются тем же законам и возбуждаются при тех же условиях, что и периферические нейроны, которые были изучены экспериментально. Этому утверждению физиолога психолог иногда противопоставляет другое мнение, а именно, что организация центрального возбуждения протекает иначе, что оно определяется рамками получаемого индивидом опыта» (Boring, 1932, р. 32). Проблема возникает особенно тогда, когда проводятся нейропсихологические эксперименты, предполагающие разрушение мозга. Результаты этих экспериментов послужили основанием для защиты двух противоположных точек зрения. По мнению одних авторов, каждая кортикальная точка, каждая клетка или группа клеток специализируется на выполнении какой-нибудь одной функции. Интеграция, необходимая для объяснения поведенческих и психологических процессов, согласно этой точке зрения, достигается за счет врожденных или образующихся при жизни прочных ассоциативных связей между нейронами. Другая точка зрения, которой придерживается и автор этой книги, состоит в том, что в мозговой ткани возникают определенные важные для организации поведения и психологического процесса взаимодействия и эти взаимодействия нельзя рассматривать только как постоянные ассоциативные связи между нейронами. К. Лешли, первый сторонник этой точки зрения, построил свою аргументацию по трем линиям: 1) эквивалентность рецепторной, функции; 2) спонтанная реорганизация моторных реакций и 3) сохранение поведения после разрушения любой части мозга, в то время как лишь разрушение всей мозговой системы устраняет раз возникшее поведение. Остановимся на этих трех ли-4 пнях доказательства. Что касается эквивалентности рецепторной функции, то эксперименты показывают, что «абсолютные характеристики стимула ямеют сравнительно небольшое значение для поведения и что реакция определяется отношениями возбуждений, которые при возникновении в любой группе рецепторных клеток этой системы имеют одинаковый эффект». Например, «животное, тренированное в выборе большего из трех кругов, может сразу же положительно реагировать на поле с наиболее широкими линиями при сравнении трех полей с полосами различной ширины» (Lashley, 1960, 238—239). Подробные данные такого рода содержатся во второй части книги; сейчас же достаточно отметить, что такая проблема существует. Что касается спонтанной моторной реорганизации, то «результаты показывают, что, когда привычно используемые двигательные органы перестают функционировать вследствие их удаления или паралича, наблюдается непосредственное спонтанное использование других моторных систем, которые ранее не были связаны •с этой деятельностью или не использовались при ее осуществлении» (Lashley, 1960, р. 239). Например, основной рисунок мышечного напряжения во время .акта письма, когда человек держит карандаш в пальцах, сохраняется и тогда, когда он переходит к письму, зажав карандаш зубами. Каким образом мозг обеспечивает это, составляет содержание первых глав третьей части этой книги. Доказательство взаимозаменяемости частей функциональных систем организма в организации поведения и психологических процессов распространяется также и на центральные отделы мозга. Это доказательство содержится во второй части. Здесь же лам необходимо только отметить, что даже значительное повреждение мозга может оказать лишь небольшое влияние на то крайне сложное поведение, которое, казалось бы, должно быть особенно чувствительно к разрушению, если считать, что интеграция целиком зависит от наличия постоянных ассоциативных связей. Лешли (см. Beach и др., 1960) следующим образом резюмирует проблему: «Эти три линии доказательства показывают, что некоторые хорошо координированные виды деятельности, казалось бы зависящие от определенных областей коры, могут в известных пределах осуществляться любой частью коры. Это может происходить в результате наличия многих дублирующих рефлекторных путей, проходящих через кору, и такое объяснение, по-видимому, можно дать всем известным случаям сохранения функций после частичного разрушения определенных областей, имеющих отношение к этим функциям. Но оно не годится для объяснения фактов сенсорной и моторной эквивалентности. Эти факты говорят о том, что, если была выработана условная реакция (например, положительная реакция на определенную структуру зрительного сигнала), эта реакция будет вызываться возбуждением сенсорных клеток, которые раньше никогда не стимулировались таким способом. Точно так же однажды приобретенные двигательные акты (например, открывание задвижки ящика) могут быть сразу же выполнены двигательными органами, которые не включались ранее в выполнение этях актов» (р. 237—240) В утверждении Лешли сметаны по крайней мере два вопроса: взаимозаменяемость частей организма и перенос навыка, который? включает в себя проблему сходства и новизны. Тем не менее приведенные здесь высказывания Лешли говорят о необходимости согласования данных психологии с данными нейрофизиологии. Такое согласование предполагает более широкое использование фактов, касающихся работы мозга, чем то, которое возможна сейчас вследствие узкого толкования нейронной теории. Между тем окончательная точка зрения на функции мозга должна быть основана на классических нейрофизиологических данных. Поэтому остановимся на тех последних успехах нейрофизиологической техники, которые показывают, что такое согласование действительно возможно (Bullock, 1959). Этот путь позволяет провести различие между первичными структурами интранейронных нервных импульсов и первичными интернейронными структурами активности; такой путь значительно расширяет диапазон тех подходов, которые применяются для изучения функций мозга. МИКРОСТРУКТУРА СОЕДИНЕНИЙ Значение нейроэлектрических процессов, генерируемых в синапсе и позади него, стало очевидным для нейрофизиологов после того, как были получены определенные данные. В течение многих лет электрическая активность, отводимая от кожи черепа, рассматривалась как отражение совокупности нервных импульсов, генерируемых нейронами мозга, лежащими где-то ниже участка записи. Когда стала возможной одновременная регистрация электрической активности локальных участков мозговой ткани и их совокупности, это предположение стало вызывать сомнение (Purpura, 1958). Запись электрической активности нейрона при внутриклеточном отведении показывает, что даже при отсутствии широко распространяющихся нервных импульсов возникают ритмические изменения медленных потенциалов, и, когда их сравнивают с ритмами, одновременно записанными от группы нейронов той же самой точки, они совпадают (Creutzfeldt et al., 1966; Fugita and Sato, 1964; Elul and Adey, 1966; Morrell, 1967). Когда мембрана нервной клетки подвергается поляризации постоянным током, амплитуда внутриклеточных ритмов меняется и это изменение отражается в суммарной записи (Eccles, 1964, см. рис. 1-4). Таким образом, было получено серьезное доказательство того, что-главный вклад в электрическую активность, записываемую суммарно в виде электроэнцефалограммы (ЭЭГ), вносится медленно возникающими флуктуациями потенциалов нейронных мембран (рис. 1-5). Только тогда, когда активность большого числа единиц синхронизирована — когда их разряды вызываются вспышкой
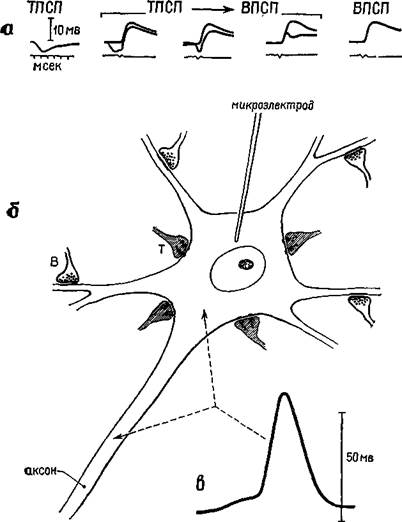
Рис. 1-4. Схема нейрона (б) с возбудительными (Б) и тормозными (Т) •синапсами и внутриклеточные записи (а) медленных постсинаптических потенциалов (ВПСП и ТПСП) — вверху. Следует отметить, что импульс, генерируемый на начальной части аксона, имеет амплитуду выше 50 мВ, тогда как амплитуда внутриклеточных записей имеет диапазон около 5 мВ. Только нервному импульсу аксона свойственно распространение (см. Eccles, 1967).
Рис. 1-5. Записи медленных дендритных потенциалов (справа) с указанием примерного уровня их регистрации в коре. Калибровка 0,2 мВ; 10 мсек (Purpura, 1967). света или звуковым шелчком, — суммарная запись и импульсы, генерируемые отдельными нейронами, оказываются одинаковыми (рис. 1-7 и 1-8). В одном таком исследовании (Fox and O'Brien,. 1965) животному предъявлялось много световых вспышек (от 3000 до 5000). Когда разряды нейрона были усреднены для всего
Рис 1-6. Диаграмма связи между внутриклеточными записями медленных потенциалов и ЭЭГ, записанной с поверхности коры той же самой области мозга, для 500-секундного периода времени (Adey, 1967).

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|