|
|
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕСОЛИТОНОВ НА ДНК Марио Салерно первым начал компьютерное экспериментирование с солитонами на ДНК не только как с формальными математическими структурами, он попытался связать их поведение в одномерном прост-ранстве полинуклеотидов с их биогенетическими, а точнее, с эпиге-нетическими функциями[18]. При этом он развил первую модель солитонов на ДНК, предложенную Инглендером и соавторами[19]. Эта модель и в последующем ее более детальные формы, включая нашу (см. ниже), представлена в понятиях механических систем как цепочка осцилляторов (оснований ДНК), связанных упругими нелинейными сахаро-фосфатными связями. Вслед за Салерно основное внимание мы уделили реально существующим известным последовательностям ДНК и влиянию их на характер поведения солитонов. На первом этапе мы повторили его эксперименты, но на существенно более длинных отрезках ДНК. Действительно, солитонные возбуждения типа кинков чувствительны к месту своей инициации, и продвижение их вдоль одной из цепочек ДНК, когда они раскрыты вследствие тепловых флуктуаций, сопровождается специфической модуляцией траектории кинков во времени. Такие солитоны являются структурами, излучающими электромагнитное и акустическое поле, их внутренняя колебательная структура способна отобразить и ретранслировать тексты и иные знаковые структуры ДНК во внутри- и внеклеточное пространство, по крайней мере на уровне крупных блоков последовательностей. В качестве примера можно привести поведение кинка на фрагменте ДНК длиной 1020 пар оснований из вируса саркомы птиц. C-район ДНК (1 (5’ начало)
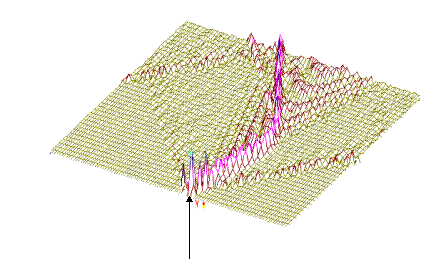
На рис.1 и рис. 2 кинки имеют форму пиков “горных гряд”, а не ступенек, поскольку взята производная от функции уравнения синусГордона. Здесь горизонтальная ось последовательность ДНК, верти-кальная амплитуда солитона. Ось на зрителя время. Видно, как при изменении места инициации солитона на определенных последо-вательностях полинуклеотида заметно меняется динамика этой уеди-ненной волны в форме ее колебательных движений вдоль цепочки ДНК. Исследуемый район молекулы богат функционально (и семантически) биологически значимыми участками, и мы вправе ожидать, что они, эти участки, будут изменять, модулировать, то есть вводить ДНК “текстовую” информацию в солитонную волну как в переносчик генетических сообщений. Такая модуляция колебательной структуры солитонов отчетливо наблюдается на приведенных графиках. Можно полагать, что спектральный состав частот колебаний солитонов является одним из механизмов преобразования текстовых структур ДНК и РНК в волновую форму и средством передачи генетических и иных сообщений в одномерном пространстве вдоль цепочек полинуклеотидов и (или) в трехмерном измерении генома как отдельной клетки, так и тканевого континуума биосистемы.
400-ый Рис.1 Влияние нуклеотидной последовательности ДНК на динамику конфор-мационного возмущения уединенной (солитоноподобной ) волны. Последо-вательность нуклеотидов - вирус саркомы птиц (первые 600 пар оснований). Центр возмущения - 400-ый нуклеотид. 450-ый Рис.2 То же, что на рис.1, но центр возмущения цепочки ДНК на 450-ом нуклеотиде. Так работает компьютерная модель динамики солитонов, в определенной мере развитая Салерно после ее выдвижения Инглендером. Салерно дал формализм, описывающий вращательные колебания нуклеотидов молекулы ДНК, для того чтобы объяснить экспериментальные данные по водородно-тритиевому обмену в ДНК. Согласно этой модели по Инглендеру, в цепи ДНК могут возникать (под воздействием теплового шума) и распространяться открытые состояния (“плавление” двойной спи-рали ДНК на коротких участках, обогащенных АТ-парами ) в виде локализованных дислокаций ( уединенных волн). Марио Салерно, про- должая работу Инглендера, в упрощенном варианте выявил влияние последовательности нуклеотидов на нелинейную динамику вращательных колебаний нуклеотидов на однотяжных участках ДНК, образующих такие открытые (“open state”) области. Позднее Якушевич, Федянин, Хомма и др. рассмотрели различные обобщения модели Инглендера, с оценкой особенностей строения ДНК, учитывая обрыв водородной связи при открытии оснований, парность цепи ДНК и другие степени свободы, отличные от вращательных. Однако, в указанных работах недостаточно сказано о причинах возникновения дислокаций в ДНК. Мы предлагаем возможный механизм этого процесса в ДНК, альтернативный гипотезе Инглендера о воздействии теплового шума как причины раскрытия пар оснований. Мы считаем, что дислокации на ДНК могут возникать при изменении периода спирали ДНК (основная часть идеи принадлежит М.Ю.Маслову). В нашей модели нуклеотиды ДНК рассматриваются как осцилляторы, подвешенные на невесомом нерастяжимом стержне; сахаро-фосфатная связь между соседними нуклеотидами в цепи моделируется линейными пружинами; спирализация вдоль цепи не учитывается; водородные связи между комплементарными основаниями моделируется “гравитационным” потенциалом. Гамильтониан по М. Салерно выглядит следующим образом: где: Коэффициенты Уравнения движения для разности
где произведена замена В случае
”непрерывный аналог” системы (2). Это уравнение имеет солитонные решения, в частности, односолитонное решение, или кинк, соответствует дислокации в цепи. Основным предположением моделей ИнглендераСалерно является то, что взаимодействие между комплементарными основаниями описывается потенциалом В нашей работе рассматривается следующий потенциал :
Кроме того, учитывается вязкость водной среды (в воде вязкость Рассматриваются также факторы, приводящие к спирализации ДНК, при этом они считаются внешними силами, задаваемыми потенциалом
где Уравнения (2) с потенциалом
Известно, что период спирали ДНК меняется в зависимости от влажности. В частности, для кристаллической ДНК В нашей работе на основе результатов численного моделирования, представленных ниже, выдвигается следующая гипотеза: изменение шага спирали может привести не только к суперспирализации, но и к локальному распариванию цепи ДНК. Кроме того, при суперспирализации напряжение в цепи снимается не полностью, поэтому локальное распаривание, вероятно, может происходить и одновременно с суперспирализацией. Система (5) численно интегрировалась в интервале
Период спирали в системе (5) Граничные условия следующие (назовем их “квазициклическими”):
Особенностью данной модели является то, что при переходе из состояния с периодом в 10 пар в состояние с периодом в 10, 5 пар почти вся цепь оказывается денатурированной (“расплавленной”). Приведенные ниже результаты описывают процесс ренатурации такой цепи с возникновением дислокаций. В этих экспериментах варьировались параметры: 1) диссипация На рис. 3 и 4 представлены результаты численного интегрирования системы (5). Показана не сама функция Можно сделать следующие выводы: 1) Способность к образованию дислокации в этой модели сильно зависит от 2) Способность к образованию дислокации также сильно зависит от параметра на рис. 1.а, 2.а ), дислокация возникла. В пользу этого утверждения также свидетельствует сравнение рис. 3(а) и 4(г). Как показывают дополнительные расчеты, влияние 3) На рис. 3(а), 4(в,г) видно, что дислокация имеет кинкообразную форму. Ширина дислокации зависит от параметров Развивая дальше модели солитонных возбуждений в ДНК (совместно с М.Ю.Масловым и др.) мы использовали условия, при которых цепочки ДНК моделируются набором ровибронных осцилляторов, подвешенных на невесомом нерастяжимом стержне; для простоты спирализация цепи не учитывается, а ровибронные степени свободы одной из цепочек считаются “замороженными”. В этом случае гамильтониан для “активной” цепочки записывается в следующем виде: H=H0+H1+H2
где:
а) а)x0=200 б)x0=250 Рис.3
в) в) x0=300 г) x0=350 Рис. 4 описывающий нелинейную связь между “активной” и “замороженной” ( При малых
имеют вид:
где произведена замена В случае
”непрерывный аналог” системы (2). Это уравнение имеет солитонные решения, в частности, односолитонное решение, или кинк, характеризующий динамику распространения дислокации в цепи. В соответствии с (1) система нелинейных уравнений движения записывается следующим образом:
Как видим, системы (2) и (4) существенно различаются. Отметим, однако, что проведенное нами численное моделирование динамики систем (2) и (4) показало следующее: если в качестве начальных условий для численного интегрирования (2) выбрать односолитонное решение его “непрерывного аналога” (3) - кинк (см. выше), то обнаруживается принципиальное сходство в характере решений. Однако, при задании начальных условий в следующем виде:
где
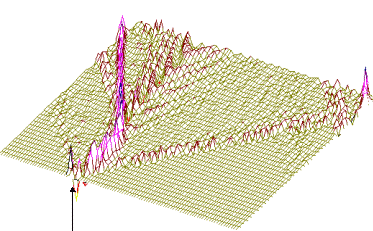
Численное интегрирование системы (2) ( рис. 1) показало, что образуются две уединенных волны, движущихся справа налево по цепи с постоянной скоростью. Первая волна имеет форму квазикинка, а вторая волна имеет форму квазибризера, причем скорость первой волны превосходит таковую для второй. Обе волны за счет “квазициклических” граничных условий, доходя до левого конца, появляются на правом конце без изменения своей формы. Квазикинк, проходя по цепи маятников, изменяет координату каждого маятника на угол На рис. 2 представлены результаты интегрирования системы (4) при тех же условиях. Из рисунка видно, что образуются те же две уединенных волны - квазикинк и квазибризер. Но принципиальное отличие от рассмотренного случая состоит в том, что квазикинк в самом начале движется с отрицательным ускорением, так что в результате его скорость оказывается меньше скорости квазибризера. Заметим, что исследования проводились на однородной поли-A-последовательности; так что изменение скорости квазикинка нельзя объяснить влиянием неоднородности цепочки. Этот эффект объясняется нелинейным взаимодействием между ее мономерами. Рис. 3 иллюстрирует результаты интегрирования системы (4) при тех же условиях за исключением того, что A=2. В данном случае реализуется только квазикинк и его отрицательное ускорение в начале движения таково, что в результате он движется в направлении, противоположном первоначальному. При интегрировании системы (2) в аналогичных условиях также образуется только квазикинк. Его скорость не меняется по сравнению со случаем рис. 1. Существенно, что при соответствующих условиях в системе типа ДНК или РНК могут возникнуть перевзбужденные ровибронные состояния. На квантовом языке это было бы адекватно перезаселению высоко лежащих квантовых уровней по сравнению с основным (реализации инверсной заселенности). В этом случае возникает заманчивая мысль, связанная с принципиальной возможностью создания биосолитонного лазера (БСЛ) на молекулах ДНК[22]. Однако, в теории динамики биополимеров хорошо известно, что конформационные движения реализуются по механизму ограниченной диффузии ввиду сильного влияния диссипативных сил со стороны микроокружения. По этой причине решение проблемы создания БСЛ на ДНК представляется весьма проблематичным, по крайней мере, для подтверждения идеи необходимо выполнение условий: Существует также и другая сложность в отношении самосогласования биосолитонов и волны электромагнитного переизлучения. Напомним, что математическое моделирование в данном случае проводилось на монотонной поли-A ДНК и поэтому оставалось неясным влияет ли гетерогенная естественная последовательность ДНК на динамику солитонного возбуждения в молекуле. Чтобы проверить это, как и ранее, был взят С-район ДНК на 3’-конце вируса саркомы птиц в качестве полигона для запуска солитонов на разных участках полимера. На этот раз вычисляли производную от функции с тем, чтобы нагляднее показать движения солитонов. На рис.5,6 (см. ниже) хорошо видно, как при сдвиге области возбуждения солитонной волны от правой нижней части графика налево траектория волны претерпевает существенные изменения, т.е. “словесно-речевое” наполнение ДНК отображается в поведении солитона. Но главное здесь не только и не столько в этом. На этот раз характерно не качание волны около некоторого положения равновесия, а движение ее в левую часть цепочки после определенного временного интервала. В этом видится определенный биологический смысл. Солитон как потенциальный “субъект чтения” ДНК должен “просматривать” протяженные контекстные зоны, а не застревать на одних и тех же “словах”.
Рис.5 а) Результаты численного моделирования динамики распространения возмущений в ДНК на основе системы (2) при значении параметра A=1. б) То же, вид сверху.
Рис.6 а) Результаты численного моделирования динамики распространения возмущений в ДНК на основе системы (4) при значении параметра A=1. б) То же, вид сверху.
Рис.7 а) Результаты численного моделирования динамики распространения возмущений в ДНК на основе системы (4) при значении параметра A=2. б) То же, вид сверху.
200-ый Рис.8 Солитонное возбуждение ДНК, но с учётом нелинейности ковалентных связей в сахаро-фосфатном остове ДНК. Последовательность нуклеотидов - вирус саркомы птиц (первые 600 пар оснований). Центр возмущения - 200-ый нуклеотид.
400-ый Рис.9 То же, что на рис. 8, но центр возмущения - 400-ый нуклеотид.
500-ый Рис.10 То же, что на рис. 9, но центр возмущения - 500-ый нуклеотид. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
 1020 нуклеотид) на 3’-конце вируса саркомы птиц. Содержит несколько “семантически” определенных участков, таких, как полипептид-кодирующий участок (между 558 и 675 нуклеотидами); PolA (936) 3’-конец вирусной РНК, сайт поли-аденилирования; 916 нуклеотид 5’-конец вирусной РНК (“capping site”); Red-участок () короткий концевой повтор вирусного генома; Pro вероятный компонент промотора транскрипции (между 870 и 900); палиндром-”шпилька” (870 912)[20].
1020 нуклеотид) на 3’-конце вируса саркомы птиц. Содержит несколько “семантически” определенных участков, таких, как полипептид-кодирующий участок (между 558 и 675 нуклеотидами); PolA (936) 3’-конец вирусной РНК, сайт поли-аденилирования; 916 нуклеотид 5’-конец вирусной РНК (“capping site”); Red-участок () короткий концевой повтор вирусного генома; Pro вероятный компонент промотора транскрипции (между 870 и 900); палиндром-”шпилька” (870 912)[20]. 1020 (3’-конец)
1020 (3’-конец)

 (1)
(1)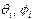 углы вращений нуклеотидов в разных цепях,
углы вращений нуклеотидов в разных цепях,  константы упругости вдоль цепей,
константы упругости вдоль цепей,  число пар в цепи,
число пар в цепи,  момент инерции оснований,
момент инерции оснований,  константа упругости водородных связей между комплементарными основаниями.
константа упругости водородных связей между комплементарными основаниями. в уравнении (1) определяются в соответствии с правилом:
в уравнении (1) определяются в соответствии с правилом:  в случае АТ и ТА пар,
в случае АТ и ТА пар,  в случае ГЦ и ЦГ пар;
в случае ГЦ и ЦГ пар;  параметр, определенный Федяниным и Якушевич[21] и полученный на основе модели синус-Гордона и экспериментальных данных. Далее для упрощения модели считается, что
параметр, определенный Федяниным и Якушевич[21] и полученный на основе модели синус-Гордона и экспериментальных данных. Далее для упрощения модели считается, что 
 , полученные из (1), имеют по М. Салерно вид:
, полученные из (1), имеют по М. Салерно вид: (2)
(2) .
. , в системе (2) можно перейти к безразмерному дифференциальному уравнению синус-Гордона:
, в системе (2) можно перейти к безразмерному дифференциальному уравнению синус-Гордона: , (3)
, (3)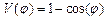 (4), в котором не учитывается обрыв водородной связи.
(4), в котором не учитывается обрыв водородной связи.
 ~ 1).
~ 1).
 период спирали.
период спирали. и с учетом вязкости принимают вид:
и с учетом вязкости принимают вид: (5)
(5) , а в водной среде
, а в водной среде  в пределах от 10. 3 до 10. 6. Именно этим фактором обусловлено явление суперспирализации. При изменении шага спирали в цепи ДНК (с фиксированными или замкнутыми концами) возникает напряжение, связанное с недостатком (избытком) количества витков спирали до релаксированного состояния. Если
в пределах от 10. 3 до 10. 6. Именно этим фактором обусловлено явление суперспирализации. При изменении шага спирали в цепи ДНК (с фиксированными или замкнутыми концами) возникает напряжение, связанное с недостатком (избытком) количества витков спирали до релаксированного состояния. Если  , то при переходе из сухого в увлажненное состояние для цепи длиной в 300 пар оснований возникнет избыток в
, то при переходе из сухого в увлажненное состояние для цепи длиной в 300 пар оснований возникнет избыток в  витка.
витка.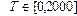 с шагом
с шагом  . Начальные условия следующие:
. Начальные условия следующие:

 длина poly(A)-цепи - 300 пар оснований. То есть параметры периода спирали в начальных условиях и в системе (5) различны. Таким образом смоделирован перенос ДНК из кристаллического состояния в увлажненное.
длина poly(A)-цепи - 300 пар оснований. То есть параметры периода спирали в начальных условиях и в системе (5) различны. Таким образом смоделирован перенос ДНК из кристаллического состояния в увлажненное.
 2) отношение параметров упругости
2) отношение параметров упругости 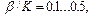 3) угол обрыва водородных связей
3) угол обрыва водородных связей 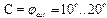 .
. , а разница
, а разница  , поскольку область изменения функции
, поскольку область изменения функции  (приблизительно от
(приблизительно от  до
до  ) велика по сравнению с характерными изменениями в системе (приблизительно от 0 до 9). Горизонтальная часть графиков соответствует нераспаренному участку цепи с периодом спирали
) велика по сравнению с характерными изменениями в системе (приблизительно от 0 до 9). Горизонтальная часть графиков соответствует нераспаренному участку цепи с периодом спирали  . Наклонная часть графиков на рис. 3(a), 4(а) соответствует дислокации.
. Наклонная часть графиков на рис. 3(a), 4(а) соответствует дислокации. . При
. При  дислокация возникла во всех рассмот-ренных случаях.
дислокация возникла во всех рассмот-ренных случаях. . Во всех случаях, когда параметр
. Во всех случаях, когда параметр  велик (
велик ( 
 на эффект проявляется в меньшей степени. Дислокация образуется или не образуется вне зависимости от значения
на эффект проявляется в меньшей степени. Дислокация образуется или не образуется вне зависимости от значения 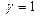 или
или 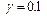 ). При больших значениях
). При больших значениях  дислокация образуется медленнее, чем при меньших.
дислокация образуется медленнее, чем при меньших. (чем больше
(чем больше  , тем меньше ширина дислокации) и
, тем меньше ширина дислокации) и 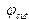 (чем больше
(чем больше  , тем меньше ширина дислокации).
, тем меньше ширина дислокации).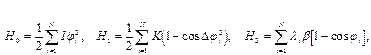 (1)
(1) - число пар оснований в цепи;
- число пар оснований в цепи; 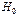 - гамильтониан, описывающий собственные осцилляции мономеров (
- гамильтониан, описывающий собственные осцилляции мономеров (  - углы вращения нуклеотидов в цепочке,
- углы вращения нуклеотидов в цепочке,  - момент инерции оснований);
- момент инерции оснований);  - гамильтониан , характеризующий нелинейно-периодическую связь между осцилляторами (
- гамильтониан , характеризующий нелинейно-периодическую связь между осцилляторами (  - константа упругости цепочки,
- константа упругости цепочки,  ),
),  - гамильтониан,
- гамильтониан, (а)
(а)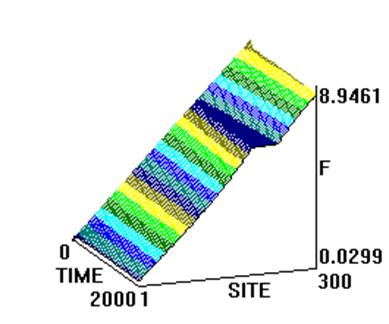 (б)
(б) б)
б) 
 в)
в)  г)
г)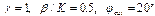 г)
г) 
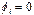 ) цепочками ДНК (
) цепочками ДНК (  - константа упругости водородных связей между комплементарными основаниями, коэффициенты
- константа упругости водородных связей между комплементарными основаниями, коэффициенты  в уравнении (1) определяются в соответствии с правилом:
в уравнении (1) определяются в соответствии с правилом: 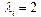 в случае АТ и ТА пар,
в случае АТ и ТА пар, 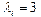 в случае ГЦ и ЦГ пар;
в случае ГЦ и ЦГ пар;  гамильтониан
гамильтониан  , что совпадает с соответствующей частью общего гамильтониана, использованного ранее (см. выше). В этом случае уравнения движения для
, что совпадает с соответствующей частью общего гамильтониана, использованного ранее (см. выше). В этом случае уравнения движения для  , полученные из (1),
, полученные из (1), (2)
(2) .
. в системе (2) можно перейти к безразмерному дифференциальному уравнению синус-Гордона:
в системе (2) можно перейти к безразмерному дифференциальному уравнению синус-Гордона: , (3)
, (3)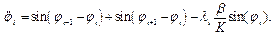 (4)
(4) (5)
(5) -”ступенчатая” функция с высотой ступени
-”ступенчатая” функция с высотой ступени  и углом наклонауступа A, выявилось различие динамики данных систем (срав. рис.1 и 2,3). Более точно, системы (2) и (4) численно интегрировались методом Рунге-Кутта четвертого порядка с начальными условиями, заданными в виде (7), в интервале
и углом наклонауступа A, выявилось различие динамики данных систем (срав. рис.1 и 2,3). Более точно, системы (2) и (4) численно интегрировались методом Рунге-Кутта четвертого порядка с начальными условиями, заданными в виде (7), в интервале  с шагом
с шагом  . Граничные условия - “квази-циклические”:
. Граничные условия - “квази-циклические”: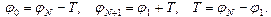
 . Варьировался параметр A (угол наклона уступа функции
. Варьировался параметр A (угол наклона уступа функции  ).
). (маятник делает полный оборот). Поэтому, проходя по замкнутой цепи маятников К раз, он изменяет координату каждого маятника на угол
(маятник делает полный оборот). Поэтому, проходя по замкнутой цепи маятников К раз, он изменяет координату каждого маятника на угол  Этим объясняется “уступообразная” форма графика на рис. 1.
Этим объясняется “уступообразная” форма графика на рис. 1. где
где  и
и  ‑ ширина и скорость солитона соответственно,
‑ ширина и скорость солитона соответственно, 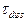 ‑ время диссипации. Положив
‑ время диссипации. Положив 
 5A и
5A и  (скорость звука), имеем оценку
(скорость звука), имеем оценку  . Отметим, что характерное время диссипации за счёт водных гидродинамических сил
. Отметим, что характерное время диссипации за счёт водных гидродинамических сил  а время затухания, обусловливаемое процессами внутри самой молекулы
а время затухания, обусловливаемое процессами внутри самой молекулы  (см., напр., Шайтан К.В. Биофизика. М.,1994. Т.39. С.949.; Чернавский и др. 1986. № 287. С. 21.).
(см., напр., Шайтан К.В. Биофизика. М.,1994. Т.39. С.949.; Чернавский и др. 1986. № 287. С. 21.). а)
а)  б)
б) а)
а) б)
б) a)
a) б)
б)