|
|
Коллоквиум по биохимии человека № 5.1. При сепсисе, травме и других метаболических стрессах распад белков становится выше синтеза (отрицательный азотистый баланс). При этом необходимо повысить количество вводимого белка до 1,3-1,5 г/кг/день, несмотря на то, что в терминальном состоянии, например, вследствие почечной или печеночной недостаточности, поступление белков следовало бы ограничить. 2. Азотистый баланс – разница между поступающим азотом в форме белка и его выведением в форме неусвоенного белка кишечником и мочевины почками. При недостаточном или неполноценном белковом питании у человека развивается отрицательный азотный баланс, т.к. поступающего белка недостаточно для возмещения потерь выводимого кишечником и почками азота. Поступление белков с пищей можно считать достаточным, если они компенсируют потери (т.е. наблюдается равновесный азотистый баланс). Для взрослых суточная доза азота около 0,8 г/кг/день. 3. Незаменимые АК: арг, вал, гис, иле, лей, лиз, мет, тре, три, фен. Арг и гис частично заменимы (незаменимы для детей). Данные АК называются незаменимыми, т.к. в организме не синтезируются углеродные скелеты этих АК и невозможно их образование с помощью реакций трансаминирования. Последствия недостаточности незаменимых АК в пище: отрицательный азотистый баланс, остановка роста и истощение, нарушения со стороны НС, неполное усвоение других АК. Коэффициент изнашивания – количество азота, теряемое человеком, находящимся на безбелковой диете (примерно 20 г белка в сутки). 4. Белки в желудке перевариваются до АК, которые затем абсорбируются клетками кишечника и попадают в портальную систему. Гидролиз белков (протеолиз) обеспечивается: 1) НСl – секретируется обкладочными кл слизистой желудка, функции: а) понижает рН химуса б) денатурирует белки, вызывает их набухание в) создает оптимальный рН для действия пепсина г) инициирует ограниченный протеолиз пепсиногена д) бактерицидные свойства 2) гастрин – гормон, секретируемый в ответ на поступление химуса в желудок, стимулирует секрецию НСl обкладочными кл и секреция пепсиногена главными клетками. 3) пепсиноген – профермент пепсина, гидролизует внутренние пептидные связи в пищевых белках 4) пепсин – преимущественно гидролизует пептидные связи, образованные ароматическими группами ароматических и больших алифатических АК с образованием больших пептидных фрагментов. 5) ренин – в желудочном соке грудных детей расщепляет белок молока – казеин. 5. Оценка кислотообразующей функций желудка имеет большое значение при диагностике язвенной болезни, гастрита, злокачественных новообразований и др. Общую кислотность желудочного сока в основном определяют: а) свободная НCl – это НСl, которая выделяется обкладочными клетками в чистом виде (20-40 ммоль/л) б) связанная с белками НCl в) кислые фосфаты г) органические кислоты. Содержание в норме 40-60 ммоль/л у взрослых, 2,8 ммоль/л у детей. Принцип определение: титрование желудочного сока с помощью р-ра NaOH в присутствии фенолфталеина и последующий расчет общей кислотности. 6. Определение содержания свободной соляной кислоты в желудочном соке: отсасывают содержимое желудка с помощью зонда, затем энтерально (пробный завтрак) или парэнтерально (гистамин) стимулируют секрецию HCl, вновь отсасывают желудочный сок и титруют его р-ром NaOH с индикатором до изменения окраски. Затем рассчитывают свободную кислотность. В норме она равна у взрослых 20-40 ммоль/л, у детей 0,5 ммоль/л. 7. Эндопептидазы – расщепляют пептидные связи внутри целой молекулы белка. Работают при оптимальных рН и концентрации электролитов. Синтезируются в виде неактивных проферментов, затем активируются путем ограниченного протеолиза. Каждая эндопептидаза специфична по отношению к определенным пептидным связям, продукт действия одного фермента может быть субстратом для другого. Основные эндопептидазы желудка и поджелудочной железы: 1) пепсин – гидролиз пептидных связей, образованных карбоксильными группами ароматических и больших алифатических АК с образованием больших пептидных фрагментов. 2) трипсин – гидролизует пептидные связи, образованные карбоксильными группами основных АК – лиз и арг. 3) химотрипсин – гидролизует пептидные связи, образованные карбоксильными группами ароматических АК (фен, тир, три) 4) эластаза – гидролизует пептидные связи, образованные карбоксильными группами маленьких алифатических АК (гли, ала, сер) 8. Протеазы ЖКТ активируются по механизму ограниченного протеолиза - избирательного гидролиза одной или нескольких пептидных связей в молекуле белка с изменением функциональной активности последнего. Ограниченный П. имеет первостепенное значение для регуляции обмена веществ в организме: 1) образование и инактивация ферментов, гормонов и других БАВ, контроль активности основных биорегуляторов (пепсиноген ® пепсин, ПОМК ® АКТГ, проинсулин ® инсулин). 2) образовании ферментов, участвующих в свертывании крови и фибринолизе 3) активация системы комплемента 4) активация ренин-ангиотензинной и калликреин-кининовой систем 5) является одним из важнейших механизмов репродукции Ограниченный П. представляет собой один из основных механизмов посттрансляционной модификации — процессинга белков. 9. Экзопептидазы– удаляют АК последовательно от N- или C-конца белковой молекулы. а) Карбоксипептидазы (вырабатываются в поджелудочной железе): карбоксипептидаза А – отщепляет нейтральные АК с С-конца пептида, карбоксипептидаза В – основные АК с С-конца пептида. б) Аминопептидазы (вырабатываются на границе тощей и подвздошной кишки): аланинаминопептидазы (отщепляют ала с N-конца), лейцинаминопептидазы (отщепляют все остальные АК с N-конца). 10. Гниение белков – совокупность превращений белков, вызванные деятельностью микроорганизмов кишечника. При гниении образуется ряд токсических продуктов распада: фенол, крезол, индол, скатол, сероводород, меркаптаны. Данные продукты попадают в портальную систему и обезвреживаются в печени путем конъюгации с глюкуроновой кислотой или с другими веществами с образованием нетоксичных и растворимых соединений – эфироглюкуроновых парных кислот. 11. Повышение активности аспарагиновой или аланиновой аминотрансфераз в крови позволяет распознавать патологические состояния, сопровождающиеся некрозом тканей, что и используется для диагностики ряда заболеваний, контроля за их течением, прогноза в будущем (пример: инфаркт миокарда, гепатит, рак печени, мышечная дистрофия). 12. Диагностическое значение определения активности аминотрансфераз в сыворотке крови: а) увеличенный уровень может свидетельствовать о: инфаркте миокарда, гепатите, алкогольном повреждении печени, раке печени, мышечной дистрофии печени. б) сниженный уровень – о пищевой недостаточности пиридоксина, при беременности, состоянии после гемодиализа. Например, при инфаркте миокарда активность АсАТ в крови резко возрастает (в 5—10 раз по сравнению с нормой) через 4—6 ч после начала заболевания, а затем постепенно снижается, достигая нормы примерно через 5 дней. Повторное повышение активности АсАТ в крови говорит о продолжающемся процессе некротического распада ткани миокарда. В клетках сердечной мышцы (миокарда) содержится гораздо больше АсТ, чем АлТ, а в клетках печени - наоборот: АлТ намного больше, чем АсТ. Поэтому ПРИ ИНФАРКТЕ МИОКАРДА АКТИВНОСТЬ АСТ в крови БУДЕТ ЗНАЧИТЕЛЬНО ВЫШЕ, ЧЕМ АЛТ, А ПРИ ВИРУСНОМ ГЕПАТИТЕ АКТИВНОСТЬ АЛТ БУДЕТ ВЫШЕ, ЧЕМ АСТ. 13. Аланинаминотрансферазы в мышцах обеспечивают удаление азота АК при их усиленном катаболизме. При повреждении мышц уровень АлАТ и АсАТ увеличивается, что может служить дополнительным диагностическим критерием. См. вопрос 11 и 12. 14. Повышенное содержание в крови АлАТ может свидетельствовать о некротических процессах в тканях (вопрос 12)
15. Аллергические реакции нередко сопровождаются падением АД, т.к. гистамин, выделяемый тучными кл, является вазодилятатором.
Образование гистамина из гистидина катализируется гистидиндекарбоксилазой.
16. Реакция образования ГАМК катализируется глутаматдекарбоксилазой в кл серого вещества головного мозга: ГАМК является медиатором тормозных импульсов в нервной системе. ГАМК и ее аналоги применяются в медицине как нейротропные средства для лечения эпилепсии и других заболеваний.
17. Пути синтеза серотонина (катализируется декарбоксилазой ароматических АК):
Роль гистамина: 1) участник аллергических реакций 2) сильный вазодилятатор 3) расширяет капилляры и увеличивает сосудистую проницаемость 4) понижает артериальное давление 5) повышает тонус (спазм) гладких мышц (бронхи) 6) усиливает секрецию желудочного сока Роль серотонина: 1) сужает сосуды 2) регулирует свертывание крови 3) обладает антиаллергическим действием. 18. Реакция синтеза дофамина (катализируется декарбоксилазой ароматических АК) и его дальнейшее использование.
Роль дофамина: 1) является предшественником катехоламинов 2) нейромедиатор 3) метил-ДОФА – сильный ингибитор декарбоксилазы ароматических АК.
19. Глутаматдегидрогеназа – играет ключевую роль в обмене АК: Глутамат-ДГ - никотинамидная, отнимаемые протоны и электроны не передаются сразу на кислород, а транспортируются по полной цепи МтО с образованием воды и параллельным образованием трех молекул АТФ. Регуляторным ферментом - ингибируется избытком АТФ, и активируется избытком АДФ.
20. Глутаматдегидрогеназная реакция (с коферментом НАДФ) – в обратном направлении:
21. Роль непрямого дезаминирования АК: 1) Обеспечивает образование новых АК из числа заменимых. 2) Образование необходимых клетке кетокислот из заменимых АК 3) Обеспечивают синтез мочевины 4) Главный путь удаления азота у АК при их интенсивном катаболизме Суть процесса: перенос аминогруппы одной АК на a-кетокислоту с образованием другой АК и другой a-кетокислоты. 22. Реакции образования новых АК: 1) переаминирование (трансаминирование) 2) прямое гидролитическое дезаминирование (асн ® асп) 3) аминирование (глу ® глн) 4) w-декарбоксилирование (асп ® ала) 23. Аммиак может накапливаться в клетках и при участии митохондриальной глутаматдегидрогеназы активировать восстановительное дезаминирование a-кетоглутаровой кислоты, элиминируя ее тем самым из цикла трикарбоновых кислот, что ведет к угнетению тканевого дыхания и накоплению кетоновых тел. Интоксикация аммиаком раньше всего проявляется симптомами угнетения ц.н.с., в тяжелых случаях может развиться кома. 24. Пути обезвреживания аммиака в организме и в клетке: а) временное связывание – в тканях, интенсивно продуцирующих аммиак – в нервной и мышечной: 1) связывание NH3 с глутаминовой, реже аспарагиновой кислотами с образованием глн и асн 2) аминирование остатков глу и асп в составе белков 3) восстановительное аминирование a-кетоглутарата в глутамат, который в реакции трансаминирования с ПВК образует ала – резервный и транспортный источник аммиака. б) общее (конечное) обезвреживание 1) выведение в виде солей аммония 2) синтез мочевины 25. У детей незаменимыми являются заменимые у взрослых АК: гис и арг. Роль гис: является предшественником важного биоамина – гистамина. Роль арг: 1) участвует в синтезе креатина в почках 2) в составе белков аргинин как полярная положительно заряженная аминокислота участвует в образовании ионных связей и в формировании гидратной оболочки белков 3) участник орнитинового цикла мочеобразования 26. Синтез мочевины – главный путь обезвреживания аммиака в печени.
27. Основными транспортными формами аммиака в печень и почки является ала и глн. 29. В случае чувствительности при некоторых формах лейкоза опухолевых клеток к недостатку аспарагина для лечения можно использовать АсАТ (аспарагиновую аминотрансферазу). 30. Другие органы, кроме почек и печени также могут обезвреживать аммиак (см. вопрос 24 – местные механизмы) 31. Значение определения мочевины в крови в клинической практике: а) повышение уровня мочевины в крови может свидетельствовать о 1) нарушении функции почек (хронической и острой почечной недостаточности), 2) внепочечные причины: обезвоживание организма, усиленный распад белков (острая желтая дистрофия печени, злокачественные опухоли и др.). б) понижение концентрации мочевины в крови: 1) повышенная скорость клубочковой фильтрации (у беременных молодых женщин, при нагрузке чрезмерным объемом внутривенных вливаний) 2) патологическое изменении значительной части паренхимы печени 3) недостаточности белка в питании, продолжительном голодании 4) врожденное нарушение нормального протекания цикла мочевины (у детей). 32. Содержание мочевины в крови в норме 2,5-8,3 ммоль/л. Причины повышения – см. выше. При уремии следует ограничить поступление белков с пищей. При этом, предпочтение отдают белковым продуктам, в которых представлены в правильном соотношении все эссенциальные аминокислоты и ограничивают при этом растительный белок, чтобы нормализовать и поддержать азотистый баланс и нейтрализовать симптоматику отравления. В случае проведения гемодиализа поступление белков ограничивать не стоит, возможно даже увеличение количества вводимых белков выше нормы. 33. Возможные пути использования безазотистых остатков АК - кетокислот в кл: 1) источник энергии (a-кетоглутаровая к-та, ЩУК ® ЦТК) 2) синтез новых АК 3) образование кетогенных тел 4) обезвреживание аммиака (глутаматдегидрогеназная реакция) Кетокислоты обеспечивают интеграцию метаболизма АК, углеводов и липидов. 34. АК могут служить источником энергии в клетке. Гликогенные АК могут превращаться в ПВК, a-кетоглутаровую к-ту, ЩУК, сукцинил-КоА, фумаровую к-ту ® ЦТК ® энергия. Кетогенные АК превращаются в ацетил- или ацетоацетил-КоА и идут на синтез кетонных тел ® энергия. 35. Избыток белков ® образование избытка гликогенных и кетогенных АК ® повышение содержания ПВК, ацетил-КоА, ацетоацетил-КоА ® синтез ВЖК и кетоновых тел и их запасание 36. Глу – гликогенная АК: а) реакция трансаминирования глу + ПВК ® a-КГ + ала б) реакции ЦТК: a-КГ ® сукцинил~КоА ® сукцинат ® фумарат ® малат ® ЩУК б) реакции глюконеогенеза: ЩУК® ФЕПВК ® ФР-1,6-ФФ ® ФР-6-Ф ® гл-6-ф ® гл 37. Из ала можно синтезировать гл: 1) реакция трансаминирования: ала + a-КГ®ПВК + гл 2) реакции глюконеогенеза: ПВК®ЩУК ® ФЕПВК ® ФР-1,6-ФФ ® ФР-6-Ф ® гл-6-ф ® гл 38. Проявления недостаточности ферментов, участвующих в обмене фенилаланина и тирозина: 1) фенилкетонурия - нарушен синтез фенилаланин-гидроксилазы, поэтому фенилаланин превращается в фенилпируват, который оказывает токсическое воздействие на развитие некоторых отделов головного мозга. 2) альбинизм - нарушен синтез ферментов, превращающих ДОФА в ДОФА-хром, поэтому нарушается синтез меланинов. 3) алкаптонурия - нарушен синтез диоксигеназы гомогентизиновой кислоты, она выделяется с мочой, моча приобретает черный цвет. 4) кретинизм - нарушен синтез йодиназы, что приводит к нарушению синтеза йодсодержащих гормонов щитовидной железы. 5) может быть нарушен синтез фермента тирозиназы, который катализирует превращение тирозина в ДОФА, следовательно будет нарушаться синтез гормонов мозгового слоя надпочечников и меланина. Из всех этих заболеваний в настоящее время удается лечить фенилкетонурию (из рациона ребенка исключают фенилаланин и увеличивают в пище количество тирозина). Если ребенка держать на этой диете до 6-7 лет, тогда не возникает умственная отсталость, т.к. к 6-7 годам успевают развиться отделы головного мозга, развитие которых задерживается при избытке в ткани мозга фенилпирувата.
39. Остаточный азот сыворотки крови – азотсодержащие небелковые вещества (промежуточные или конечные продукты обмена простых и сложных белков) – мочевина, мочевая кислота, креатин, креатинин, аммиак, индикан, билирубин и т.д. Азот этих веществ называют остаточным, т.к. он остается в фильтрате после осаждения белков. Основная часть остаточного азота крови – азот мочевины (50%), азот АК (25%) и азот других азотсодержащих соединений. Норма остаточного азота крови для взрослых 14,3-25,0 ммоль/л. Остаточный азот определяют в безбелковом фильтрате после осаждения белков с последующей минерализацием фильтрата концентрированной серной кислотой. При этом образуется сульфат аммония, который образует с реактивом Несслера соединение желто-оранжевого цвета. Интенсивность окрашивания пропорциональна содержанию азота. Значение метода: диагностика поражения почек (исследование выделительной функции) и мочевинообразовательной функции печени. 40. Нуклеотиды – мономеры нуклеиновых кислот – состоят из: 1) азотистого основания (у всех нуклеиновых кислот) 2) пентозы (рибозы у РНК или дезоксирибозы у ДНК) 3) остатка фосфорной кислоты Азотистое основание + пентоза = нуклеозид. Свойства нуклеотидов: 1) отрицательно заряжены (за счет фосфатных групп) 2) циклические соединения 3) гидрофобны 4) поглощают свет при 260 нм (УФ область). Функции нуклеотидов: 1) структурная – мономеры нуклеиновых кислот, входят в состав коферментов 2) энергетическая (АТФ - это универсальный аккумулятор энергии, энергия УТФ используется для синтеза гликогена, ЦТФ - для синтеза липидов, ГТФ - для движения рибосом в ходе трансляции (биосинтез белка) и передачи гормонального сигнала (G-белок) 3) регуляторная - аллостерические эффекторы многих ключевых ферментов, цАМФ и цГМФ являются посредниками в передаче гормонального сигнала при действии многих гормонов на клетку и активаторами протеинкиназы 41. Первичная структура НК – линейная последовательность нуклеотидов в одной цепи. Нуклеотиды связываются через остатки фосфорной кислоты с помощью 3’,5’-фосфодиэфирных связей.
42. Вторичная структура ДНК – пространственная ориентация полинуклеотидных цепей в ее молекуле. Представляет собой двойную правозакрученную спираль диаметром 1,8-2,0 нм. Полинуклеотидные цепи антипараллельны и комплементарны. Двойную спираль стабилизируют: 1) водородные связи между азотистыми основаниями 2) Вандерваальсовы силы между азотистыми основаниями 3) гидрофобные взаимодействия 43. Принцип в основе формирования двойной спирали в молекуле ДНК – комплементарность (А-Т (две водородные связи), Г-Ц(три водородные связи). В РНК такой принцип не реализуется, т.к. она одноцепочечная, в гибридах типа ДНК-РНК комплементарность реализуется в процессе транскрипции, при этом Г соответствует Ц, а А – У. 44. Третичная структура ДНК и тРНКнеобходима для компактности молекул в ядре (ДНК) и в цитоплазме (тРНК). Третичная структура тРНК формируется самостоятельно (в виде двойной буквы Г), а ДНК – в результате связи с белками с образованием нуклеопротеинов 4-х уровней упаковки: нуклеосомный, соленоидный, петлевой, уровень метафазной хромосомы. 45. Нуклеосома – глобула (октамер), состоящая из белкового ядра (из 8 молекул-гистонов, Н2А, Н2В, Н3, Н4 – по две молекулы каждого вида), вокруг которого ДНК делает 1,5-2 оборота. При этом длина накрученного фрагмента ДНК порядка 50 нм, а компактизация составляет в 5-7 раз по сравнению с исходной. Гистоновые белки обладают положительным зарядом в связи с большим содержанием основных АК – арг и лиз, связь между белками и ДНК – ионная. 46. Нуклеопротеины под действием желудочного сока (НCl) распадаются на белки и нуклеиновые кислоты. Белки катаболизируются до АК, затем всасываются, НК – до нуклеозидов, которые также всасываются. В кл с ними происходит распад либо до АО и пентозы, либо до АО и фосфопентозы:
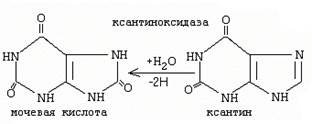
47. При гиперурикемии повышается образование мочевой кислоты под действием ксантиноксидазы:
48. Причины повышения уровня мочевой кислоты в крови: 1) нечувствительность к регуляторам ферментов синтеза пуринов de novo 2) снижение активности ферментов реутилизации пуринов 3) почечная патология 4) нехватка витамина B9 Последствия: гиперурикемия и подагра (накопление кристаллов мочевой кислоты в суставах). 49. Синтез пуриновых нуклеотидов путем: а) повторного использования готовых азотистых оснований (характерно для размножающихся тканей) б) de novo из низкомолекулярных предшественников (источники N – аспартат, глицин, глутамин, источники С – СО2, глицин, двухуглеродные фрагменты метенил-ТГФК и формил-ТГФК).
ФРПФ + глутамин -------> глутамат + ФФ + фосфорибозиламин (катализируется ключевым ферментом фосфорибозиламидотрансферазой)
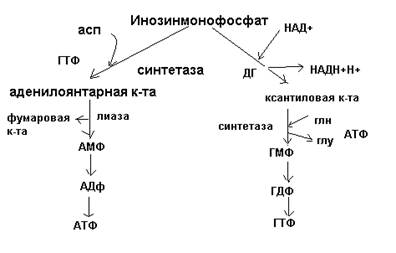
Основные промежуточные продукты: фосфорибозиламин, инозинмонофосфат, аденилоянтарная, ксантиловая кислоты. 50. Синтез пиримидиновых нуклеотидов. Сначала образуется циклическая структура пиримидинового азотистого основания, и только затем присоединяется рибозо-5-фосфат. Источники атомов – СО2, аспартат, глутамин.
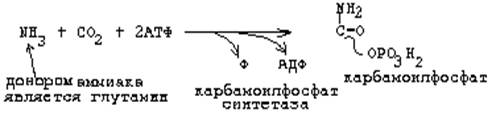
Карбамоилфосфатсинтетаза II является ключевым ферментом.
Основные промежуточные продукты: карбомоилфосфат, оротовая к-та, дигидрооротовая к-та. 51. Дефицит фолиевой кислоты (витамина В9) может привести к нарушению синтеза пуринов и пиримидинов ® нарушение синтеза НК, процессов репарации. 52. Реакция, ведущая к образование карбамоилфосфата, общая для синтеза пиримидиновых оснований и мочевины: а) для пиримидинов (карбамоилфосфатсинтетаза II):
б) для мочевины:
53. Дезоксирибонуклеотиды образуются из рибонуклеотидов путем восстановления рибозного остатка при участии специфической фосфатной системы и фермента рибонуклеозидредуктазы. Донором ионов водорода в этой реакции служит тиоредоксин – низкомолекулярный белок, содержащий SH-группы.
Тиоредоксин получает два атома водорода от трипептида глутатиона, переходящего при этом в окисленную форму. Последующее восстановление окисленного глутатиона происходит с помощью фермента глутатионредуктазы, использующей для этого НАДФН2. 54. Репликация – процесс удвоения ДНК, происходящий в S-фазу клеточного цикла. Полуконсервативный процесс. Главный фермент – ДНК-полимераза, ведущая синтез дочерней цепи по принципам комплементарности, антипараллельности, в одном направлении от 5’ к 3’ концу. Субстраты для синтеза ДНК: нуклеозид-3-фосфаты: дАТФ, дГТФ, дЦТФ, дТТФ. ДНК-полимераза также требует кроме субстратов, ионы магния и праймер. Праймер – затравочный олигонуклеотид, синтезируемый праймазой. Репликация начинается в местах повышенной концентрации пар А-Т. Фермент хеликаза раскручивает двойную спираль, фермент топоизомераза снимает напряжение в области репликативной вилки и предотвращает обратное скручивание, ДНК-полимераза I удаляет остатки праймера. Терминация репликации наступает у человека после 8-9 часов, при этом ДНК-лигаза сшивает отдельные фрагменты Оказаки. 55. Репарация – внутриклеточный процесс восстановления поврежденной из-за неблагоприятных воздействий структуры ДНК. Различают: а) дорепликативную б) репликативную в) пострепликативную репарации. Прямая репарация – химическая реакция, направленная на восстановление структуры поврежденного нуклеотида, эксцизионная репарация – вырезание поврежденного фрагмента с участием ряда ферментов: 1) эндонуклеазы – узнает поврежденный участок и разрывает рядом с ним нить ДНК 2) экзонуклеазы – вырезает поврежденный участок 3) ДНК-полимеразы – комплементарно достраивает фрагмент ДНК на месте разрушенного 4) лигаза – сшивает концы ресинтезированного участка с основной нитью ДНК. Роль репарации: обеспечение постоянства генетического материала. 56. Синтез и-РНК. – транскрипция – переписывание информации с ДНК на и-РНК. Для эукариот в этом процессе участвуют: РНК-полимераза I – синтезирует р-РНК, РНК-полимераза II – синтезирует и-РНК, РНК-полимераза III – т-РНК. Субстраты синтеза: матрица, рибонуклеозидтрифосфаты, праймер не требуется. Синтез идет антипараллельно, от 5’ к 3‘ концу. Вначале и-РНК образуется в виде предшественника – пре-и-РНК, затем идет кэпирование – присоединение 7-метилгуанозина к 5’-концу полиаденилового конца для защиты этого конца от нуклеаз, помогает присоединиться к рибосомам. На 3’- конце идет полиаденилирование – для защиты от нуклеаз, участвует в транспорте из ядра в цитоплазму. 57. Единица транскрипции у эукариот – транскриптон: неинформативная зона (промотор) и информативная (структурные гены – экзоны – информативны, интроны – нет + спейсеры – вставки, разделяющие структурные гены). В отличие от прокариот у эукариот работу транскриптона регулируют несколько генов-репрессоров, а индукторами являются сложные молекулы (гормоны и т.д.), часто требуется несколько индукторов – ступенчатый процесс. Образуемая в процессе транскрипции и-РНК содержит неинформативные участки и подвергается процессингу (вырезание неинформативных участков) и сплайсингу (склеиванию оставшихся фрагментов). 58. Обратная транскрипция – передача генетической информации от и-РНК к ДНК с помощью фермента обратной транскриптазы (ревертазы). В ходе этого процесса комплементарно синтезируется одна цепь ДНК на основе и-РНК, а затем достраивается вторая цепь ДНК на основе первой. Роль: 1) один из способов репарации ДНК 2) получение множественных копий ДНК 3) передача генетической информации у ряда вирусов 59. Генетический код – система записи генетической информации в ДНК (РНК) в виде определенной последовательности нуклеотидов. Последовательность нуклеотидов определяет последовательность включения АК в синтезируемый белок. 3 нуклеотида – триплет – кодон – кодируют 1 АК. Совокупность триплетов и составляет генетический код. Свойства генетического кода: 1) триплетность 2) вырожденность (избыточность – 1 АК кодируется несколькими триплетами) 3) специфичность (1 кодон кодирует только 1 АК) 4) однонаправленность (от 5’ к 3’ концу) 5) неперекрываемость (один нуклеотид входит в состав только одного кодона) 6) универсальность (у всех живых организмов одинаковые АК кодируются одинаковыми кодонами) 7) отсутствие знаков препинания внутри гена. 60. тРНК (10-15% от все РНК) транспортирует АК к месту синтеза белка – на рибосомы. Имеет форму клеверного листа, содержит два основных центра: а) антикодон – для связывания АК б) участок для прикрепления АК (5’ и 3‘ концы) Нуклеотидная цепь тРНК содержит всего 75—90 нуклеотидов. Особенностью тРНК является относительно высокое содержание нуклеотидов, включающих минорные азотистые основания. Т-РНК с помощью высокоспецифичных ферментов аминоацил-тРНК-синтетаз присоединяют к себе ту или иную аминокислоту и переносят ее на рибосому. Для одной и той же аминокислоты имеется несколько тРНК, которые называют изоакцепторными. Транспортная РНК в ходе синтеза полипептидной цепи белка «узнает» специфическую аминоацил-тРНК-синтетазу, принимает от нее активированную аминокислоту, присоединяется к иРНК на рибосоме и тем самым обеспечивает строгую специфичность выбора и встраивания аминокислот в растущую молекулу белка; после образования пептидной связи между доставленной аминокислотой и уже построенной полипептидной цепью тРНК удерживает эту цепь на рибосоме. 61. Аминоацил-тРНК-синтетаза – фермент, участвующий в биосинтезе белка, отвечает за специфичность связывания АК с тРНК. АК + АТФ ® ФФн + аминоацил-аденилат + тРНК ® АМФ + аминоацил-тРНК. Известно 20 аминоацил-тРНК-синтетаз (по количеству L-АК). Содержит 4 участка: для АК, для т-РНК, для воды, для АТФ. Правильность присоединения АК. 62. Рекогниция – процесс узнавания тРНК своей АК, происходящий при помощи фермента аминоацил-тРНК-синтетазы. 63. Трансляция – синтез полипептида с последовательностью АК, отвечающей последовательности триплетов нуклеотидов в молекуле и-РНК. Этапы трансляции: 1) кодирование 2) рекогниция 3) собственно трансляция: инициация, элонгация, терминация. Участники трансляции: и-РНК, рибосомы, аминоацилтРНК, активированные формы АК, ГТФ, факторы инициации, элонгации, терминации. Перед началом синтеза белка рибосомы диссоциированы. На этапе инициации к малой субъединицы рибосомы 3’ концом присоединяется и-РНК. К первому инициирующему кодону присоединяется 1 аминоацил-тРНК, несущая метионин. Затем присоединяется к образованному комплексу большая субъединица рибосомы с затратой ГТФ – образование инициирующего комплекса. В рибосоме образуется два центра – аминоацильный и пептидильный. В свободный А-центра поступает следующая аминоацил-тРНК ® фермент пептидил трансфераза образует пептидную связь между АК, дипептид остается в А-центре® транслоказа перемещает рибосому на 1 кодон и-РНК ® дипептид при этом оказывается в П-центре ® в освободившийся А-центр поступает следующая АК. Элонгация заканчивается терминацией – наступает, когда в А-центр оказывается один из трех нонсенс-кодонов (стоп-кодонов): УАА, УАГ, УГА. Они не соответствуют ни одной тРНК, распознаются рилизинг-ферментами, которые вызывают отсоединение синтезированного белка и диссоциацию рибосомы. 61. Регуляция транскрипции у прокариот. Единица транскрипции прокариот – оперон, состоит из 1) промотора – места первичного прикрепления РНК-полимеразы, 2) оператора – включает и выключает экспрессию структурных генов 3) структурных генов. На некотором расстоянии от оперона находится ген-регулятор, отвечающий за синтез белка-репрессора, способного вступать в химическое взаимодействие с геном-оператором и «выключать» его работу. Поступление в кл индуктора ® индуктор+белок-репрессор ® освобождение гена-оператора ® РНК-полимераза присоединяется к промотору ® комплементарно со структурных генов создается и-РНК ® трансляция и-РНК ® синтез фермента, разрушающего индуктор ® освобождение белка-репрессора ® присоединение его к оператору ® остановка трансляции. 65,67. ПЦР – увеличение количества копий ДНК, находящейся в биологическом материале в минимальных количествах. Этапы: 1) денатурация ДНК (до 90°) 2) добавление специфического праймера и охлаждение ДНК (до 55°) (отжиг) 3) добавление нуклеотидов (субстратов синтеза) и ДНК-полимеразы (фермента синтеза) 4) повторение цикла. Позволяет за короткое время получить множество копий ДНК (порядка 20 млн за 0,5 ч). Применение: 1) диагностика вирусных и бактериальных инфекций (ВИЧ, гепатит, хламидиоз) 2) генетическая диагностика заболеваний 3) судебная медицина. 66. Расшифровав нуклеотидную последовательность гена, можно установить последовательность АК в кодируемом белке, однако следует помнить что ген содержит некодирующие последовательности – интроны и спейсеры, которые удаляются в ходе процессинга и сплайсинга и не отражаются в структуре конечного белка. Поэтому, чтобы достоверно установить последовательность АК в белке, можно методами генетической инженерии синтезировать белок, а затем подвергнуть его биохимическому анализу (электрофорезу). 68. Клонирование – получение большого количества молекул, клеток, организмов – потомков одного предка. Для клонирования бактериальных и вирусных генов необходимы носители генетической информации – плазмиды, ДНК (РНК) бактериофага, нуклеоид, хромосомы дрожжей. Этапы: 1) получение генетического материала 2) включение гена в векторную молекулу, создание рекомбинантной ДНК 3) введение рекДНК в кл хозяина 4) отбор трансформированных кл на селективных средах. Применение: 1) получение разнообразных вакцин и иммунологических диагностикумов 2) синтез ряда БАВ: СТ, инсулин, эритропоэтин, интерфероны, факторы свертывания крови 3) получение ферментов в промышленности 4) получение относительно недорогого пищевого белка для животных и т.д. 69. Вектор – природный ген определенного микроорганизма с внедренным в него участком чужеродного гена. В качестве векторов применяется: 1) плазмида – небольшая кольцевидная молекула ДНК бактерий, реплицируемая независимо от нуклеоида 2) бактериофаг лямбда 3) хромосомы дрожжей 4) космидные векторы - гибрид фага лямбда и плазмиды. 70. «Метод отпечатков пальцев» ДНК используется для: 1) идентификации личности в судебной медицине 2) комплексной диагностики наследственных заболеваний 3) поиске участков ДНК, отвечающих за развитие патологии 4) установления отцовства. В основе метода – принцип гибридизации на основе комплементарности. 71. Рестриктазы (рестриктационные эндонуклеазы) – ферменты, узнающие специфическую последовательность нуклеотидов и разрывающие в этом месте молекулу ДНК. Действуют в области палиндромов ДНК – мест, где последовательность нуклеотидов одной цепи идентична последовательности нуклеотидов другой цепи, прочитанной в обратном порядке. В молекулярной биологии рестриктазы используют для создания генетических векторов (внедрение гена в плазмиду и т.п.) 72. При проведении блот-анализа ДНК используют одноцепочечную молекулу ДНК, комплементарную искомой и меченую радиоактивным фосфором P32. Позволяет диагностировать в геноме дефектные гены, установить наличие наследственных заболеваний. 73. Этапы постановки блот-анализа ДНК: 1) экстракция ДНК 2) разрезание ДНК рестриктазами 3) разделение ДНК путем электрофореза в агарозном геле 4) перенос ДНК на нитроцеллюлозу и денатурация ее щелочью 5) блокирование пустых зон избытком ДНК 6) обработка зондом и образование гибридов мишень-зонд 7) отмывка несвязанного зонда и ауторадиография 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2024 stydopedia.ru Все материалы защищены законодательством РФ.
|