
|
|
Пиридинзависимые дегидрогеназыК числу пиридинзависимых дегидрогеназ относится свыше 150 ферментов, которые катализируют восстановление НАД и НАДФ различными органическими субстратами. Структура НАД была выяснена Эйлером и соавт. Варбургу принадлежит заслуга открытия НАДФ. НАД и НАДФ являются динуклеотидами, в которых мононуклеотиды связаны между собой через остатки фосфорной кислоты. В состав одного из нуклеотидов входит амид никотиновой кислоты (витамин РР), другой представляет собой адениловую кислоту. В молекуле НАДФ имеется еще один остаток фосфорной кислоты, присоединенный к рибозе в положении С3'. Катализируемые пиридинзависимыми дегидрогеназами реакции можно изобразить так:
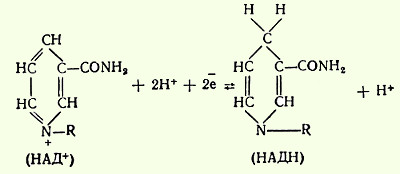
Способность НАД и НАДФ играть роль промежуточного переносчика водорода связана с наличием в их структуре амида никотиновой кислоты. В электронно-протонной форме обратимое гидрирование - дегидрирование (присоединение и отдача протонов и электронов) этих коферментов может быть представлено с помощью следующего уравнения
При переносе двух восстановительных эквивалентов от субстрата на НАД+ один из них затем присутствует в восстановленном коферменте в виде водорода, а другой - в виде электрона. Свободный ион Н+ остается в среде. Для простоты обычно восстановленные формы НАД и НАДФ изображают символами НАДН2 и НАДФН2. Окисленные и восстановленные пиридиннуклеотиды обладают характерными спектрами поглощения в УФ-области. В окисленной форме НАД (НАДФ) имеет одну узкую полосу поглощения с максимумом при 260 нм, зависящую от наличия аденина в его структуре. В восстановленной форме (НАДН2) поглощение света в этой зоне понижается и появляется вторая широкая полоса поглощения с максимумом при 340 нм. Вторая полоса обусловлена исчезновением одной двойной связи в никотинамидном комплексе кофермента при его восстановлении. Это свойство данных коферментов позволяет использовать спектрофотометрические методы анализа для быстрого количественного определения ряда субстратов - доноров водорода, а также для исследования активности НАДФ-зависимых дегидрогеназ. В клетках НАД-зависимые дегидрогеназы преимущественно участвуют в процессах, связанных с переносом электронов от органических субстратов к кислороду. В свою очередь НАДФ-зависимые дегидрогеназы играют существенную роль в реакциях биосинтеза (например, высших жирных кислот, сте-ринов и др.). В соответствии с этим коферменты НАД и НАДФ различаются по своей внутриклеточной локализации: НАД концентрируется главным образом в митохондриях, а бoльшая часть НАДФ находится в цитоплазме клеток.
Флавиновые ферменты Следующим акцептором атомов водорода является группа флавиновых ферментов, которые осуществляют перенос водородов (протонов и электронов) от восстановленных НАД или НАДФ:
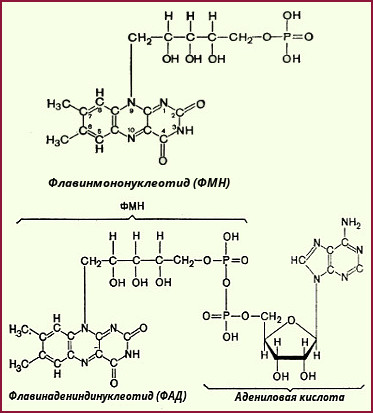
В некоторых случаях (например, при окислении сукцината или жирных кислот) флавиновые ферменты могут играть роль первичных дегидрогеназ, т. е. прямо, без участия НАД- или НАДФ-зависимых дегидрогеназ, принимать электроны и протоны от окисляемых субстратов. Насчитывается около 30 флавиновых ферментов. Это сложные белки, простетической группой которых, как уже отмечалось, являются либо ФМН, либо ФАД. ФМН и ФАД очень прочно, в отличие от коферментов НАД и НАДФ, присоединены к ферменту (точнее, к апоферменту) и не отщепляются от него ни на одной из стадий каталитического цикла. Флавиновые дегидрогеназы, входящие в систему дыхательных катализаторов животных клеток, обычно содержат в качестве простетической группы не ФМН, а ФАД. Впервые ФМН и ФАД были выделены Варбургом в середине 30-х годов XX столетия. В это же время Куном был идентифицирован витамин B2 (рибофлавин), который входит в структуру ФАД и ФМН.
Окисленные формы ФМН и ФАД обладают характерным спектром поглощения с максимумом при 450 нм; в окисленном состоянии флавиновые ферменты окрашены в красный, коричневый или зеленый цвет. При восстановлении ФАД и ФМН полоса 450 нм в спектре поглощения исчезает. Активной частью молекулы ФАД или ФМН является изоаллоксазиновое кольцо рибофлавина, к атомам азота которого могут присоединяться два атома водорода, т. е. два электрона и два протона при соответствующей внутримолекулярной перегруппировке двойных связей. Установлено, что многие из флавиновых дегидрогеназ являются сложными олигомерными образованиями, которые состоят из нескольких белковых субъединиц и содержат, помимо флавиннуклеотидов (ФАД или ФМН), также атомы металлов - чаще всего негемовое (негеминовое) железо. Это так называемые металлофлавопротеиды. По-видимому, наличие атомов металла (Fe, Мn, Сu), способных легко изменять свою валентность, присоединяя или теряя электроны, придает металло-флавопротеидам своеобразный характер, сближая их с типичными электронотранспортазами - цитохромами.
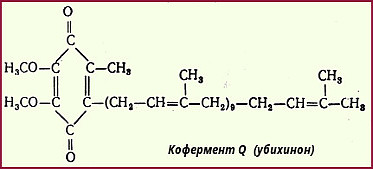
Кофермент Q (убихинон) Следующим компонентом дыхательной цепи является кофермент Q (KoQ), или убихинон. Этот кофермент впервые был описан Мортоном, который показал, что данное соединение имеет повсеместное распространение в клетках, и назвал его убихиноном. Позже было установлено, что убихинон, подобно НАД и ФАД, может играть роль промежуточного переносчика водородных атомов, т. е. электронов и протонов. KoQ является производным бензохинона с длинной боковой цепью, которая в большинстве тканей млекопитающих состоит из 10 изопреноидных единиц (KoQ10). Считают, что восстановленная форма флавиновых ферментов в цепи дыхательных катализаторов передает атомы водорода (протоны и электроны) KoQ. Как всякий хинон, KoQ может существовать как в окисленной, так и восстановленной форме:
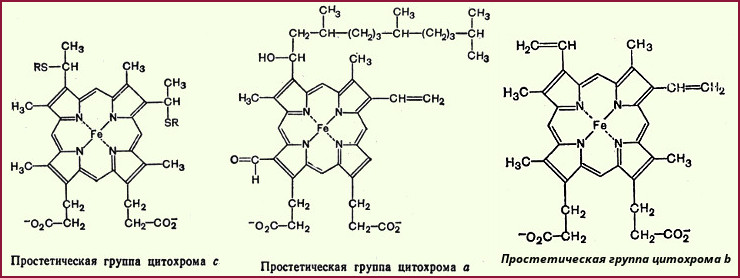
Цитохромы Дальнейший перенос электронов1 (1Принято считать, что цитохромы переносят только электроны.) от KoQ (или от восстановленной формы флавинового фермента) на кислород осуществляет система цитохромов. Данная система состоит из ряда гемсодержащих белков (гемопротеидов), открытых в 1886 г. Мак-Мунном. Однако роль этих гемопротеидов в дыхании клеток была выяснена лишь в 1925 г. Кейлином. В процессе тканевого дыхания наиболее важную роль играют цитохромы b, c1, с, аа3. Все они имеют простетическую геминовую группу, близкую к тему гемоглобина (простетическая группа цитохрома b, как считают, идентична гему). Приводим структуру простетических групп некоторых цитохромов:
Цитохромы отличаются друг от друга не только своими простетическими группами, но и белковыми компонентами. В ходе каталитического процесса валентность содержащегося в цитохромах железа обратимо изменяется (Fe2+ <--> Fe3+). Все цитохромы, особенно в восстановленной форме, имеют характерные спектры поглощения. Величины окислительно-восстановительного потенциала у разных цитохромов также неодинаковы (табл. 24). Например, для цитохрома "с" окислительно-восстановительный потенциал составляет +0,25 В, а для цитохрома "а" +0,29 В. Система 1/2O2/H2O имеет потенциал + 0,82 В. Таким образом, было выяснено, что звено цитохромов располагается в цепи окисления между убихиноном и кислородом; при этом цитохромы в цепь дыхательных катализаторов включаются в определенной последовательности: цит.b --> цит.c1 --> цит.c --> цит.аа3 Цитохромы b, c1 и с выполняют функцию промежуточных переносчиков электронов, а цитохром аа3 (цитохромоксидаза) является терминальным дыхательным ферментом, непосредственно взаимодействующим с кислородом. Окисленная форма цитохромоксидазы (Fe3+) принимает электроны от восстановленного цитохрома с, переходя в восстановленную форму (Fe2+), которая затем вновь окисляется в Fe3+-форму молекулярным кислородом. Образовавшийся "активный" кислород присоединяет два протона из окружающей среды, в результате чего и образуется молекула воды. В последнее время установлено, что цитохромоксидаза состоит из 6 субъединиц; каждая из них содержит геминовую группу и атом меди. По-видимому, две субъединицы из шести составляют цитохром а, а остальные четыре относятся к цитохрому а3.
Как уже отмечалось, иногда цепь биологического окисления может быть укорочена. Имеются флавиновые ферменты, например сукцинатдегидрогеназа, которые способны отнимать водород непосредственно от субстрата. В ряде случаев цепь дыхательного катализатора может иметь еще более простое строение. Так, например, оксидазы L- и D-аминокислот (за исключением глутаматдегидрогеназы), ксантиноксидаза и некоторые другие ферменты катализируют окисление соответствующих субстратов без участия как НАД или НАДФ, так и цитохромов. Эти оксидазы по своей химической природе являются флавопротеидами. Все ферменты тканевого дыхания - компоненты цепи дыхательных катализаторов, как установлено, связаны главным образом с митохондриями, точнее с их внутренними мембранами. Никотинамиддинуклеотидные коферменты и некоторые ферменты цикла трикарбоновых кислот "вмонтированы" в белковый слой мембраны, металлофлавопрогеиды, убихинон и цитохромы связаны с липидными ее структурами. Ферменты этого класса катализируют окислительно-восстановительные реакции, лежащие в основе биологического окисления. Класс насчитывает 22 подкласса. Коферментами этого класса являются НАД, НАДФ, ФАД, ФМН, убихинон, глутатион, липоевая кислота. Примером подклассов могут служить ферменты, действующие на СН-ОН-группу доноров, на СH-СН-группу доноров, на СН-NН2-группу доноров, на гемсодержащие доноры. Наиболее распространены следующие рабочие названия оксидоредуктаз: 1. Дегидрогеназы– оксидоредуктазы, катализирующие дегидрирование субстрата с использованием в качестве акцептора водорода любых молекул, кроме кислорода. 2.Если перенос водорода от молекулы донора трудно доказуем, то такие оксидоредуктазы называютредуктазами. 3. Оксидазы– оксидоредуктазы, катализирующие окисление субстратов с молекулярным кислородом в качестве акцептора электронов без включения кислорода в молекулу субстрата. 4. Монооксигеназы– оксидоредуктазы, катализирующие внедрение одного атома кислорода в молекулу субстрата с молекулярным кислородом в качестве донора кислорода. 5. Диоксигеназы– оксидоредуктазы, катализирующие внедрение 2 атомов кислорода в молекулу субстрата с молекулярным кислородом в качестве донора кислорода. 6. Пероксидазы– оксидоредуктазы, катализирующие реакции с пероксидом водорода в качестве акцептора электронов. Систематическое название образуется: Донор электронов : акцептор электронов – оксидоредуктаза. Первичные акцепторы водорода окислительно-восстановительных реакций относят к 2 типам дегидрогеназ: никотинамвдзависимым, содержащим в качестве коферментов производные никотиновой кислоты, и флавинзависимым, содержащим производные рибофлавина. Как уже было сказано, для проявления каталитической активности большинству ферментов необходимо наличие кофермента. Кофермент, локализуясь в каталитическом участке активного центра, принимает непосредственное участие в химической реакции, выступая в качестве акцептора и донора химических группировок, атомов, электронов. Кофермент может быть связан с белковой частью молекулы ковалентными и нековалентными связями. В первом случае он называется простетической группой (пр: FAD, FMN, биотин). Во втором случае кофермент взаимодействует с ферментом только на время химической реакции и может рассматриваться в качестве второго субстрата. Примеры - NAD+, NADP+. Никотинамидзависимые дегидрогеназы содержат в качестве коферментов NAD+ или NADP+,NAD+и NADP+ - производные витамина PP. Эти коферменты входят в состав активных центров дегидрогеназ, но могут обратимо диссоциировать из комплекса с апоферментами и включаются в состав фермента в ходе реакции. Субстраты NAD- и NADP-зависимых дегидрогеназ находятся в матриксе митохондрий и в цитозоле. Рабочей частью никотинамидных коферментов служит никотинамид . Большинство дегидрогеназ, поставляющих электроны в ЦПЭ, содержат NAD+. Они катализируют реакции типа: R-CHOH-R1 + NAD+↔ R-CO-R1 + NADH + Н+. Т.о. NAD+, присоединяя протоны и электроны от различных субстратов, служит главным коллектором энергии окисляемых веществ и главным источником электронов, обладающих высоким энергетическим потенциалом, для ЦПЭ. NADPH не является непосредственным донором электронов в ЦПЭ, а используется почти исключительно в восстановительных биосинтезах. Однако возможно включение электронов с NADPH в ЦПЭ благодаря действию пиридиннуклеотид трансгидрогеназы, катализирующей р-ию: NADPH + NAD+ ↔ NADP++ NADH. Флавиновые дегидрогеназы содержат в качестве коферментов FAD или FMN. Эти коферменты образуются в организме человека из витамина В2. Флавиновые коферменты прочно связаны с апоферментами. Рабочей частью FAD и FMN служит изоаллоксазиновая сопряжённая циклическая система. FAD служит акцептором электронов от многих субстратов в реакциях типа: R-CH2-CH2-R1 + Е (FAD) ↔R-CH=CH-R1 + Е (FADH2), где Е - белковая часть фермента. Большинство FAD-зависимых дегидрогеназ - растворимые белки, локализованные в матриксе митохондрий. Исключение составляет сукцинат-дегидрогеназа, находящаяся во внутренней мембране митохондрий. К FMN-содержащим ферментам принадлежит NADH-дегидрогеназа, которая также локализована во внутренней мембране митохондрий; она окисляет NADH, образующийся в митохондриальном матриксе. 12. АТФ - молекула, богатая энергией, поскольку она содержит две фосфоаншдридные связи (β, γ). При гидролизе концевой фосфоангидридной связи АТФ превращается в АДФ и ортофосфат Рi Величина свободной энергии гидролиза АТФ делает возможным его образование из АДФ за счёт переноса фосфатного остатка от таких высокоэнергетических фосфатов, как, например, фосфоенолпируват. В свою очередь, АТФ может участвовать в таких эндергонических реакциях, как фосфорилирование глюкозы или глицерина. АТФ выступает в роли донора энергии в эндергонических реакциях многих анаболических процессов. Наконец, за счёт свободной энергии АТФ совершаются различные виды работы, лежащие в основе жизнедеятельности организма, например, такие как мышечное сокращение или активный транспорт веществ. Таким образом, АТФ - главный, непосредственно используемый донор свободной энергии в биологических системах. В клетке молекула АТФ расходуется в течение одной минуты после её образования. У человека количество АТФ, равное массе тела, образуется и разрушается каждые 24 ч. Основным способом получения АТФ в клетке является окислительное фосфорилирование, протекающее в структурах внутренней мембраны митохондрий. При этом энергия атомов водорода молекул НАДН и ФАДН2, образованных в гликолизе, ЦТК, окислении жирных кислот, преобразуется в энергию связей АТФ. Однако также есть другой способ фосфорилирования АДФ до АТФ – субстратное фосфорилирование, поскольку он является частью метаболического пути («субстратной цепи»). Их особенностью является катализирование растворимыми ферментами. Этот способ связан с передачей макроэргического фосфата или энергии макроэргической связи какого-либо вещества (субстрата) на АДФ. К таким веществам относятся метаболиты гликолиза (1,3-дифосфоглицериновая кислота, фосфоенолпируват), цикла трикарбоновых кислот (сукцинил-SКоА) и креатинфосфат. Энергия гидролиза их макроэргической связи выше, чем 7,3 ккал/моль в АТФ, и роль указанных веществ сводится к использованию этой энергии для фосфорилирования молекулы АДФ до АТФ. Отличия: разные источники энергии, для окислительного необходимы движение электронов в дых цепи, для субстратного необ-ма энергия макроэргической связи. Существует три основных способа использования АТФ: 1. биосинтез веществ, 2. транспорт веществ через мембраны, 3. изменение формы клетки и ее движение. Эти процессы вкупе с процессом образования АТФ получили название АТФ-цикл: Использование АТФ как источника Е возможно только при условии непрерывного синтеза АТФ из АДФ за счёт Е окисления орг-их соединений. Цикл АТФ-АДФ - основной механизм обмена энергии в биологических системах. Свободное окисление. Одна из задач свободного (несопряженного) окисления – превращения природных или неприродных субстратов, называемых в этом случае ксенобиотиками (ксено – несовместимый, биос – жизнь). Они осуществляются ферментами диоксигеназами и монооксигеназами. Окисление протекает при участии специализированных цитохромов, локализованных чаще всего в эндоплазматическом ретикулуме, поэтому иногда этот процесс называют микросомальным окислением. В реакциях свободного окисления участвуют также кислород и восстановленные дыхательные переносчики (чаще всего НАДФН). Акцептором электронов является цитохром Р-450 (иногда цитохром b5). Окисление субстрата протекает по следующей схеме: SH + O2–> SOH. Ферменты свободного окисления: оксидазы, оксигеназы, некоторые дегидрогеназы. Механизм действия оксигеназ включает изменение валентности входящих в их состав ионов двухвалентных металлов. Диоксигеназы присоединяют к субстрату молекулярный кислород, активируя его за счет электрона атома железа в активном центре. Монооксигеназы требуют участия в реакции НАДФН, атомы Н2 которого взаимодействуют с одним из атомов О2, поскольку только один электрон связывается с субстратом. В процессе свободного окисления вследствие особенностей используемых цепей передачи электронов не происходит образования АТФ; биологическая роль этих процессов заключается в метаболизме ряда природных и ксенобиотических субстратов. В последнем случае свободное окисление выполняет важную функцию модификации чужеродных соединений. К последним относятся лекарственные средства, гербициды, продукты загрязнения окружающей среды, в возрастающем количестве попадающие в организм с водой, пищей и атмосферным воздухом. Как правило, они имеют гидрофобные св-ва. Многие из них являются канцерогенными. Их гидроксилирование в ходе свободного окисления облегчает последующую деструкцию и выведение из организма. При свободном окислении высвобождающаяся при сопряженном с окислением распаде химических связей энергия переходит в тепловую и рассеивается. По типу свободного окисления идут все без исключения оксигеназные реакции, все окислительные реакции, ускоряемые пероксидазами или сопровождающиеся образованием Н202, многие реакции, катализируемые оксидазами. Процессы свободного окисления сосредоточены в цитозоле, в мембранах эндоплазматической сети клетки, в мембранах лизосом, пероксисом и аппарата Гольджи, на внешних мембранах митохондрий и хлоропластов. Они идут также в ядерном аппарате клетки. Тканевые и возрастные особенности окислительных процессов. Анаэробные ткани могут получать энергию без кислорода. Такими тканями являются: скелетные мышцы, эритроциты, периферические нервы, мозговое вещество почек, кость, хрящ, соединительная ткань. Аэробные ткани получают энергию с использованием кислорода и полностью зависят от кровотока. К таким тканям относятся: головной мозг, сетчатка глаза, сердце, кора почек, печень, слизистая тонкого кишечника. Потребление кислорода, а значит, и интенсивность окислительных процессов с возрастом падают.
13. Сопряжение в дыхательной цепи— это такое состояние, когда окисление (перенос электронов) сопровождается фосфорилированием, то есть синтезом АТФ.Разобщение— это такое состояние дыхательной цепи, когда окисление идет, а фосфорилирование не происходит, то есть пункты фосфорилирования выключены полностью или частично. В этом случае вся или какая-то часть образующейся энергии выделяется в виде тепла. Разобщение в дыхательной цепи могут вызывать липофиль-ные вещества, которые способны переносить протоны водорода с внешней стороны внутренней мембраны митохондрий на внутреннюю, минуя АТФ-синтетазу. В результате вся энергия мембранного потенциала будет рассеиваться в виде тепла. Разобщение вызывают: 2,4-ДНФ (динитрофенол), многие яды промышленных производств, бактериальные токсины, набухание митохондрий, жирные кислоты, ионофоры (вещества, переносящие ионы через мембрану). Разобщители повышают скорость переноса электронов по дыхательной цепи и выводят ее из под контроля АТФ. Вещества, которые снижают величину электрохимического градиента, приводя к увеличению скорости движения электронов по ферментам дыхательной цепи, уменьшению синтеза АТФ и возрастанию катаболизма называются разобщители. Они разобщают (разъединяют) процессы окисления и фосфорилирования внутренней мембраны митохондрий. К разобщителям в первую очередь относят "протонофоры" – вещества переносящие ионы водорода. При этом одновременно уменьшаются оба компонента электрохимического градиента – электрический и химический, и энергия градиента рассеивается в виде тепла. Следствием эффекта протонофоров является возрастание катаболизма жиров и углеводов в клетке и во всем организме. Классическим протонофором является динитрофенол, жирорастворимое соединение, присоединяющие ионы водорода на внешней поверхности внутренней митохондриальной мембраны и отдающие их на внутренней поверхности. Белок термогенин является физиологическим протонофором. Кроме динитрофенола и термогенина протонофорами, к примеру, являются салицилаты, жирные кислоты и трийодтиронин. яд веществ может ингибировать ферменты дыхательной цепи и блокировать движение электронов от НАДН и ФАДН2 на кислород. Они называются ингибиторы. В результате прекращается движение электронов, выкачивание ионов Н+ и работа АТФ-синтазы. Синтез АТФ отсутствует и клетка погибает. Выделяют три основных группы ингибиторов: 1. действующие на I комплекс, например, амитал (производное барбитуровой кислоты), ротенон, прогестерон, 2. действующие на III комплекс, например, экспериментальный антибиотик антимицин А, 3. действующие на IV комплекс, например, сероводород (H2S), угарный газ (СО), цианиды (-CN). 14. Энергетическую ценность и выгодность окисления вещества можно подсчитать. Количество запасенной энергии при окислении того или иного соединения характеризует количество АТФ, а эффективность использования энергии вещества и ее захвата – коэффициент P/O.Коэффициент Р/О– это коэффициент сопряжения окисления и фосфорилирования. Это количество моль АТФ, синтезируемых из АДФ и Н3РО4 на каждый полноценный грамм-атом поглощенного О2. Для субстратов пируват, изоцитрат, малат окисляющимися Над-зависимыми ДГ коэффициент Р/О=3. Для сукцината (ФАД-зависимая ДГ) =2. Если Р/О для пирувата=3 – митохондрии находятся в сопряженном состоянии. Работа дыхательных ферментов регулируется с помощью эффекта, который получил название дыхательный контроль. Дыхательный контроль– это прямое влияние электрохимического градиента на скорость движения электронов по дыхательной цепи (т.е. на величину дыхания). В свою очередь, величина градиента напрямую зависит от соотношения АТФ и АДФ, количественная сумма которых в клетке практически постоянна ([АТФ] + [АДФ] = const). Реакции катаболизма направлены на поддержание постоянно высокого уровня АТФ и низкого АДФ. Возрастание протонного градиента возникает при снижении количества АДФ и накоплении АТФ (состояние покоя), т.е. когда АТФ-синтаза лишена своего субстрата и ионы Н+ не проникают в матрикс митохондрии. При этом ингибирующее влияние градиента усиливается и продвижение электронов по цепи замедляется. Ферментные комплексы остаются в восстановленном состоянии. Следствием является уменьшение окисления НАДН и ФАДН2 на I и II комплексах и замедление катаболизма в клетке. Снижение протонного градиента возникает при исчерпании резервов АТФ и избытке АДФ , т.е. при работе клетки. В этом случае активно работает АТФ-синтаза и через канал Fо проходят в матрикс ионы Н+. При этом градиент, естественно, снижается, поток электронов возрастает, в результате повышается выкачивание ионов Н+ в межмембранное пространство и снова их быстрое "проваливание" через АТФ-синтазу внутрь митохондрий с синтезом АТФ. Ферментные комплексы I и II усиливают окисление НАДН и ФАДН2 (как источников электронов) и снимается ингибирующее влияние НАДН на цикл лимонной кислоты и пируватдегидрогеназный комплекс. Как итог – активируются реакции катаболизма углеводов и жиров. В клетке часто может создаваться ситуация, когда реакции окислительного фосфорилирования идут с определенными вариациями. Эти вариации могут являться следствием нарушений в организме или физиологической реакцией на воздействие. 15. Дыхательный контроль Окисление субстратов и фосфорилирование АДФ в митохондриях прочно сопряжены. Скорость использования АТФ регулирует скорость потока электронов в ЦПЭ. Если АТФ не используется и его концентрация в клетках возрастает, то прекращается и поток электронов к кислороду. С другой стороны, расход АТФ и превращение его в АДФ увеличивает окисление субстратов и поглощение кислорода. Зависимость интенсивности дыхания митохондрий от концентрации АДФ называют дыхательным контролем. Механизм дыхательного контроля характеризуется высокой точностью и имеет важное значение, так как в результате его действия скорость синтеза АТФ соответствует потребностям клетки в энергии. Запасов АТФ в клетке не существует. Относительные концентрации АТФ/АДФ в тканях изменяются в узких пределах, в то время как потребление энергии клеткой, т.е. частота оборотов цикла АТФ и АДФ, может меняться в десятки раз. Общее содержание АТФ в организме 30-50 г, но каждая молекула АТФ в клетке "живёт" меньше минуты. В сутки у человека синтезируется 40-60 кг АТФ и столько же распадается. Увеличение концентрации АДФ немедленно приводит к ускорению дыхания и фосфорилирования. Разобщение дыхания и фосфорилирования Некоторые химические вещества (протонофоры) могут переносить протоны или другие ионы (ионофоры) из межмембранного пространства через мембрану в матрикс, минуя протонные каналы АТФ-синтазы. В результате этого исчезает электрохимический потенциал и прекращается синтез АТФ. Это явление называют разобщением дыхания и фосфорилирования. В результате разобщения количество АТФ снижается, а АДФ увеличивается. В этом случае скорость окисления NADH и FADH2возрастает, возрастает и количество поглощённого кислорода, но энергия выделяется в виде теплоты, и коэффициент Р/О резко снижается. Как правило, разобщители - липофильные вещества, легко проходящие через липидный слой мембраны. Одно из таких веществ - 2,4-динитрофенол, легко переходящий из ионизированной формы в неионизированную, присоединяя протон в межмембранном пространстве и перенося его в матрикс. Примерами разобщителей могут быть также некоторые лекарства, например дикумарол - антикоагулянт или метаболиты, которые образуются в организме, билирубин - продукт катаболизма тема, тироксин - гормон щитовидной железы. Все эти вещества проявляют разобщающее действие только при их высокой концентрации. Терморегуляторная функция ЦПЭ На синтез молекул АТФ расходуется примерно 40-45% всей энергии электронов, переносимых по ЦПЭ, приблизительно 25% тратится на работу по переносу веществ через мембрану. Остальная часть энергии рассеивается в виде теплоты и используется теплокровными животными на поддержание температуры тела. Кроме того, дополнительное образование теплоты может происходить при разобщении дыхания и фосфорилирования. Разобщение окислительного фосфорилирования может быть биологически полезным. Оно позволяет генерировать тепло для поддержания температуры тела у новорождённых, у зимнес-пящих животных и у всех млекопитающих в процессе адаптации к холоду. У новорождённых, а также зимнеспящих животных существует особая ткань, специализирующаяся на теплопродукции посредством разобщения дыхания и фосфорилирования - бурый жир. Бурый жир содержит много митохондрий. В мембране митохондрий имеется большой избыток дыхательных ферментов по сравнению с АТФ-синтазой. Около 10% всех белков приходится на так называемый разобщающий белок (РБ-1) - термогенин. Бурый жир имеется у новорождённых, но его практически нет у взрослого человека. В последние годы появились факты, свидетельствующие о существовании в митохондриях разных органов и тканей млекопитающих разобщающих белков, похожих по своей структуре на РБ-1 бурой жировой ткани. По своей структуре термогенин близок к АТФ/АДФ-антипортеру, но не способен к транспорту нуклеотидов, хотя сохранил способность переносить анионы жирных кислот, служащих разобщителями. На внешней стороне мембраны анион жирной кислоты присоединяет протон и в таком виде пересекает мембрану; на внутренней стороне мембраны диссоциирует, отдавая протон в матрикс и тем самым снижает протонный градиент. Образующийся анион возвращается на наружную сторону мембраны с помощью АТФ/ АДФ-антипортера. При охлаждении стимулируется освобождение норадреналина из окончаний симпатических нервов. В результате происходят активация липазы в жировой ткани и мобилизация жира из жировых депо. Образующиеся свободные жирные кислоты служат не только "топливом", но и важнейшим регулятором разобщения дыхания и фосфорилирования.
16. Митохондрии-микроскопические двумембранные полуавтономные органоиды общего назначения, обеспечивающие клетку энергией, получаемой благодаря процессам окисления и запасаемой в видефосфатных связей АТФ. Митохондрии также участвуют в биосинтезе стероидов, окислении жирных кислот и синтезе нуклеиновых кислот. Присутствуют во всех эукариотических клетках. В прокариотических клетках митохондрий нет, их функцию выполняют мезосомы - впячивания наружной цитоплазматической мембраны внутрь клетки. Митохондрии могут иметь эллиптическую, сферическую, палочковидную, нитевидную и др. формы, которые могут изменяться в течение определенного времени. Количество митохондрий в клетках, выполняющих различные функции, варьирует в широких пределах - от 50 и достигая в наиболее активных клетках 500-5000. Их больше там, где интенсивны синтетические процессы (печень) или велики затраты энергии (мышечные клетки). В клетках печени (гепатоцитах) их число составляет 800. а занимаемый ими объем равен примерно 20% объема цитоплазмы. Размеры митохондрий составляют от 0,2 до 1-2 мкм в диаметре и от 2 до 5-7 (10) мкм в длину. На светооптическом уровне митохондрии выявляются в цитоплазме специальными методами и имеют вид мелких зерен и нитей (что обусловило их название - от греч. mitos- нить иchondros- зерно). В цитоплазме митохондрии могут располагаться диффузно, однако обычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл) органелл движения (аксонем спермия, ресничек), компонентов синтетического аппарата (цистерн ЭПС). Согласно одной из гипотез, все митохондрии клетки связаны друг с другом и образуют трехмерную сеть. Митохондрия окружена двумя мембранами - наружной и внутренней, разделенныхмежмембранным пространством, и содержатмитохондриальный матрикс, в который обращены складки внутренней мембраны -кристы. · Наружная митохондриальная мембранагладкая, по химическому составу сходна с наружной цитоплазматической мембраной и обладает высокой проницаемостью для молекул массой до 10 килодальтон, проникающих из цитозоля в межмембранное пространство. По своему составу она похожа на плазмалемму, 25% составляют белки, 75% липиды. Среди липидов присутствует холестерол. Наружная мембранаа содержит много молекул специализированныхтранспортных белков (например,поринов), которые формируют широкие гидрофильные каналы и обеспечивают ее высокую проницаемость, а также небольшое количествоферментных систем. На ней находятсярецепторы, распознающие белки, которые переносятся через обемитохондриальные мембраны в особых точках их контакта -зонах слипания. · Внутренняя мембрана имеет выросты внутрь- гребни или кристы, делящие матрикс митохондрии на отсеки. Кристы увеличивают площадь поверхности внутренней мембраны. Таким образом, внутреняя митохондриальная мембрана по площади превосходит наружную. Кристы расположены перпендикулярно или продольно длине митохондрии. Кристы по форме могут быть везикулярные, тубулярные или ламеллярные. Химический состав внутренней мембраны митохондрий сходен с мембранами прокариот (например, в ней присутствует особый липид - кардиодипин и отсутствует холестерол). Во внутренней митохондриальной мембране преобладают белки, составляющие 75%. Во внутреннюю мембрану встроены белки трех типов (а) белки электрон-транспортной цепи (дыхательной цепи) - НАД'Н-дегидрогеназа и ФАД'Н дегидрогеназа - и другие транспортные белки, (б)грибовидные тельца АТФ-синтетазы (головки которых обращены в сторону матрикса) и (в)часть ферментов цикла Кребса (сукцинатдегидрогеназа). Внутренняя митохондриальная мембрана отличается чрезвычайно низкой проницаемостью, транспорт веществ осуществляется через контактные сайты. Низкая проницаемость внутренней мембраны для мелких ионов из-за высокого содержания фосфолипида Митохондрии- полуавтономные органоиды клетки, т.к. содержат собственную ДНК, полуавтономную систему репликации, транскрипции и собственный белоксинтезируюший аппарат - полуавтономную систему трансляции (рибосомы 70S типа и т-РНК). Благодаря этому митохондрии синтезируют часть собственных белков. Митохондрии могут делиться независимо от деления клетки. Если из клетки удалить все митохондрии, то новые в ней не появятся. Согласно теории эндосимбиоза митохондрии произошли от аэробных прокариотических клеток, которые попали в клетку хозяина, но не переварились, вступили на путь глубокого симбиоза и постепенно, утратив автономность, превратились в митохондрии. Митохондрии - полуавтономные органоиды, что выражается следующими признаками: 1) наличие собственного генетического материала (нити ДНК), что позволяет осуществлять синтез белка, а также позволяет самостоятельно делиться независимо от клетки; 2) наличие двойной мембраны; 3) пластиды и митохондрии способны синтезировать АТФ (для хлоропластов источник энергии - свет, в митохондриях АТФ образуется в результате окисления органических веществ). Функции митохондрий: 1) Энергетическая- синтез АТФ (отсюда эти органоиды и получили название «энергетических станций клетки»): При аэробном дыхание на кристах происходит окислительное фосфорилирование (образование АТФ из АДФ и неорганического фосфата за счет энергии, освободившейся при окислении органических веществ) и перенос электронов по электрон-транспортной цепи. На внутренней мембране митохондрии расположены ферменты, участвующие в клеточном дыхании; 2) участие в биосинтеземногих соединений (в митохондриях синтезируются некоторые аминокислоты, стероиды (стероидогенез), синтезируется часть собственных белков), а также накопление ионов (Са2+), гликопротеидов, белков, липидов; 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2024 stydopedia.ru Все материалы защищены законодательством РФ.
|
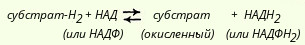
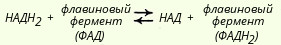


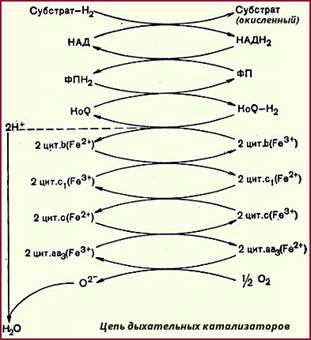 Обычно считается, что на участке между НАД и KoQ осуществляется двухэлектронный перенос, а на участке между цитохромом b и кислородом - одноэлектронный (следовательно, на каждом этапе дыхательной цени должно участвовать по два цитохрома); тем не менее нельзя исключить и другие возможности. Учитывая, что любой компонент дыхательной цепи может восстанавливать редокс-системы только с более высоким потенциалом, а окислять - с более низким (см. табл. 24), то в целом цепь дыхательных катализаторов может быть представлена следующим образом:
Обычно считается, что на участке между НАД и KoQ осуществляется двухэлектронный перенос, а на участке между цитохромом b и кислородом - одноэлектронный (следовательно, на каждом этапе дыхательной цени должно участвовать по два цитохрома); тем не менее нельзя исключить и другие возможности. Учитывая, что любой компонент дыхательной цепи может восстанавливать редокс-системы только с более высоким потенциалом, а окислять - с более низким (см. табл. 24), то в целом цепь дыхательных катализаторов может быть представлена следующим образом: