
|
|
Молекулярная динамика индивидуальных полимерных цепей, их конформации и гибкостьВ общем случае молекулярное тепловое движение в решающей степени определяет временное поведение полимерных цепей при внешнем воздействии или изменении термодинамических параметров состояния (температуры, давления, состава или объема): скорость реакции или отклонения на изменившиеся условия и скорость возникновения или рассасывания флуктуаций. В данном разделе обобщены современные молекулярно-кинетические представления об основных типах молекулярного теплового движения полимерных цепей и подходы к описанию их взаимосвязи с релаксационными процессами. При этом очевидно, что многообразие уровней молекулярной структуры полимерных цепей должно обусловливать многообразие форм и масштабов теплового движения в них. Очевидно также, что при переходе от индивидуальных полимерных цепей к полимерным телам необходимо учитывать тот факт, что решающее влияние на механизм и кинетику молекулярного теплового движения в полимерах оказывают не только их молекулярное строение, но и фазовое состояние и надмолекулярная структура, определяющие характер упаковки цепей и возможности реализации тех или иных типов молекулярного теплового движения. Универсальной формой теплового движения, проявляющейся во всех конденсированных системах, в том числе полимерных, являются мелкомасштабные безактивационные (резонансные) высокочастотные колебания атомов и групп относительно первичных (ковалентных) и вторичных (ван-дер-ваальсовских) связей с частотами, лежащими в области высокочастотного инфракрасного спектра, т.е. с периодом колебаний порядка 10-13-10-14 с. Колебания относительно ковалентных связей, подразделяемые на валентные (вдоль направления связи), деформационные (с изменением угла связи) и крутильные (вокруг направления связи), относятся к «жестким». Колебания относительно ван-дер-ваальсовских связей являются «мягкими». Высокочастотные «жесткие» и «мягкие» колебания не играют важной роли в релаксационных процессах в полимерах, т.к. характерные времена (длительность или период) внешних воздействий значительно превышают характерные времена этих колебаний. Например, минимальная длительность механического воздействия лежит в области В подавляющем большинстве случаев релаксационные процессы в полимерах в решающей степени связаны с проявлением крупномасштабных низкочастотных активационных форм теплового движения полимерных цепей: переходами повторяющихся звеньев, их сочетаний (сегментов) или отдельных частей (групп) между равновесными состояниями (потенциальными ямами) вследствие поворота относительно гибкой ковалентной σ-связи с преодолением потенциального (активационного) барьера ΔЕ при разности энергии в соседних потенциальных ямах Δе. К наиболее важным видам такого движения относятся: крупномасштабные формы сегментального движения - кооперативное и индивидуальное движение сегментов и отдельных звеньев цепей (α и β формы соответственно), а также некоторые вторичные формы сегментальной подвижности - движение отдельных концевых и боковых звеньев или их частей (γ и δ формы соответственно). В анализе динамики крупномасштабных низкочастотных активационных форм сегиентального движения в полимерах и их взаимосвязи с различными релаксационными процессами используются два подхода: локальный и глобальный. В локальном подходе используется классическая молекулярная динамика применительно к полимерным цепям: преимущественно рассматриваются различные по характеру и масштабу (интенсивности и частоте) виды внутримолекулярного (внутрицепного, или микроброуновского) ориентационно-вращательного движения отдельных структурных элементов цепей любой топологии, не сопровождающегося смещением центров масс цепей как целого. Важнейшими характеристиками молекулярной динамики при этом являются коэффициенты самодиффузии соответствующих структурных элементов и молекулярное время корреляции, или микроскопическое время релаксации как среднее время между столкновениями структурных элементов, совершающих тепловое движение, или время, за которое затухают их корреляционные функции. Это время непосредственно связано с макроскопическим временем релаксации – характерным временем соответствующего релаксационного процесса. Для молекулярно-кинетического описания низкочастотных и квазистатических процессов диэлектрической и, особенно механической релаксации, более эффективен и широко используется глобальный подход. Этот подход базируется на анализе обобщенных моделей полимерных цепей, обычно с одной крупномасштабной формой сегментального движения, которая определяет как внутримолекулярные конформационные превращения цепей и, соответственно, их гибкость, так и их диффузионную (макроброуновскую) подвижность как целого (само- и взаимодиффузию). В глобальном подходе способность полимерных цепей принимать определенные устойчивые (равновесные) конформации в результате сегментального движения характеризует их статическую (равновесную, или термодинамическую) гибкость и обратную ей величину - жесткость, а скорость или характерное время процессов изменения конформаций – их кинетическую, или динамическую гибкость и жесткость соответственно. При этом для описания закономерностей процессов изменения конформаций цепей и оценки их термодинамической (равновесной) и кинетической гибкости предложены три обобщенных статистических модели полимерных цепей: свободно-сочлененную, поворотно-изомерную и персистентную (рис.З). Модель свободно-сочлененной цепи представляет собой сочетание шарнирно связанных жестких звеньев заданной длины, совершающих свободное движение в шарнирах друг относительно друга без фиксированных углов. Статистический анализ формы пространственного клубка, принимаемого такой цепью в результате случайных поворотов звеньев в шарнирах, дает простую зависимость наиболее вероятного (равновесного) расстояния между концами цепи
Такая модель характеризует идеально гибкую цепь, сегментальное движение в которой не ограничивается пространственными (стерическими) и энергетическими факторами. Вводимые при этом относительные параметры гибкости (жесткости) применимы к сравнительно гибким (термодинамически и кинетически) полимерным цепям, содержащим большое количество σ-связей. Одним из таких показателей равновесной, или термодинамической жесткости полимерных цепей служит отношение экспериментально определенного расстояния между концами цепей в равновесных условиях (
Ряс.1. 3. Обобщенные статистические модели полимерных цепей: 1 - свободно-сочлененная; 2 - поворотно-изомерная; 3 – персистентная (l – длина звена, A –длина статистического сегмента,
Показателями равновесной, или термодинамической жесткости полимерных цепей служат также длина статистического сегмента Куна ( A= где Нижеприведены примеры значений Z, А и NА для полимерных цепей различнойжесткости, определенные по размерам статистических клубков линейных макромолекул в растворах:
ПолимерыZ А, нм NА Гибкоцепные: Полидиметилсилоксан 1,4-1,6 1,40 4,9 Цис-полиизопрен (натуральный каучук) 1,7 - - Полибудтадиен 1,7 - - Полиамид 66 2,1 - 6,6 Полиметилметакрилат 2,2 1,51 6,0 Полиизобутилен 2,2 1,83 7,3 Поливинилацетат 2,1-2,3 - 6,9 Полистирол 2,2-2,4 2,00 7,9 Полиэтилен 2,3-2,4 2,08 8,3 Поливинилхлорид 2,8 2,96 11,7 Жескоцепные: Производные целлюлозы 4,0-4,5 10,0-25,0 - Ароматический сложный полиэфир - 10,0 - (поли-п-оксибензоат) Ароматические полиамиды- 40-210 В поворотно-изомерной модели полимерной цепи ее участки (сегменты), способны скачком (с преодолением потенциального барьера) изменять свою конформацию по отношению к соседним участкам и обратно с определенной вероятностьюв результате поворота частей сегментов (звеньев) относительно На Рис.1.4 приведена общая схема нагружения полимерного клубка и сегмента полимерной цепи, состоящего из четырех атомов углерода, соединенных σ-связями, в наиболее вероятной «транс»-конформации (в форме плоского зигзага), обладающей наименьшей потенциальной энергией. Пунктиром показан поворот элементарного звена, состоящего из двух атомов, соединенных σ-связями, относительно соседней связи на угол φ. На Рис.1.4в приведена зависимость потенциальной энергии участка цепи от угла поворота. Конформации сегмента цепи при повороте элементарного звена на Статистические расчеты показывают, что длина сегмента в такой модели цепи А связана с величиной Δе экспоненциальной зависимостью:
где l0 - длина элементарного звена полимерной цепи, совершающего «транс» - «гош» переход.
Поведение цепи, состоящей из таких сегментов, рассматривается как идеально гибкое, поэтому при числе N таких сегментов средне-квадратичное расстояние между концами цепи в условиях термодинамического равновесия равно:
При Δе<<kT А→1,5 l0 и уравнение (1.65) практически совпадает с уравнением (1.1). Это свидетельствует о том, размер молекулярного клубка подчиняется степенному скейлинговому закону (закону масштабной инвариантности), означающему, что при удвоении длины цепи размер клубка увеличится в 21/2 раза. В общем случае этот закон записывается в виде:
а)
б)
в)
Рис.1.4. Схематическое изображение нагружения полимерного клубка силой f (а), сегмента, состоящего из четырех атомов углерода (б), и зависимости потенциальной энергии сегмента от угла поворота φ (в) в поворотно-изомерной модели полимерной цепи.
где ν – универсальный показатель степени, одинаковый для всех клубков в заданных термодинамических условиях и заданной размерности пространства. Так в трехмерном пространстве для идеально гибких цепей ν=1/2, а для любых цепей в «хорошем» растворителе ν=3/5. Коэффициент пропорциональности К зависит от деталей структуры звеньев цепи и условий, в которых она находится. При обобщенном анализе процессов изменения конформаций цепи как целого в поворотно-изомерной модели (подход Флори) цепь представляется в виде последовательности прямолинейных участков, состоящих из звеньев в виде плоских транс-конформеров и разделенных случайно блуждающими изломами на угол В отличиеот свободно-сочлененной и поворотно-изомерной моделей, описывающих гибкость открытых полимерных цепей, образованных преимущественно
Для характеристики термодинамической жесткости жестких полимерных цепей используются также отношение длины статистического сегмента,(удвоенной персистентной длины) к диаметру или толщине цепи: коэффициент анизометрии, или характеристическое отношение сегментов p, и доля ароматических звеньев ( Равновесные, или статические конформации, которые могут принимать полимерные цепи в различных состояниях, в глобальном подходе подразделяются на нерегулярные (статистические) и регулярные - складчатые и выпрямленные, или палочкообразные (рис.1.5). Нерегулярные конформации полимерных цепей в форме статистических клубков (см. рис.1.5, схема I) существуют при неупорядоченном распределении цепей в аморфном состоянии и в растворе, а регулярные (рис.1.5. схемы 2 - 3) - при упорядоченной упаковке цепей в кристаллических и жидко- кристаллических состояниях.
Рис.1.5. Схема равновесных конформаций полимерных цепей: 1 - статистический клубок; 2 - складчатая конформация; 3 - выпрямленная, или палочкообразная конформация ( Возможность образования регулярных равновесных конформаций цепей определяется их стереорегулярностью и гибкостью. Регулярные складчатые конформации могут образовывать только стереорегулярные гибкие цепи, универсальный критерий гибкости которых Важнейшее значении с точки зрения изучения молекулярной подвижности в гетерогенных (композиционных) полимерных материалах, в тонких полимерных пленках и покрытиях имеет влияние жестких стенок или твердых поверхностей на статическую гибкость (равновесные конформации) и кинетику процессов изменения конформаций полимерных цепей. При контакте полимерной цепи с твердой поверхностью последняя оказывает чисто стерическое (пространственное) ограничение, и в равновесных условиях это должно приводить к уменьшению конформационного набора цепи, т. е. к уменьшению ее энтропии на величину ΔS, пропорциональную (для поворотно-изомерной модели) квадрату отношения средне-квадратичного расстояния между концами цепи в плоскости
Вторым фактором, влияющим на равновесную конформацию цепи при ее контакте с твердой поверхностью, является ее притяжение (адсорбция звеньев), уменьшающее энтальпию цепи в изобарно-изотермических условиях на величину:
где N- общее число звеньев в цепи, fb–доля адсорбированных звеньев, пропорциональная отношению l0 /d0, δ – параметр, характеризующий энергию взаимодействия звеньев цепи с твердой поверхностью. Таким образом, равновесная конформация полимерной цепи при контакте с твердой поверхностью будет определяться конкуренцией между действием адсорбционных сил, увеличивающих плотность клубка, и уменьшением энтропии из-за стерических ограничений, уменьшающих плотность клубка. Очевидно, что эти эффекты должны сказываться аналогичным образом и на кинетике процесса установления равновесной конформации. Для обобщенного описания равновесных конформаций полимерных цепей и процессов их установления в равновесном состоянии при внешних воздействиях, в первую очередь механической силы и напряженности электрического поля, и соответствующих им смещениях (деформации и поляризации) в глобальном подходе используются молекулярно-кинетические методы. Пример молекулярно-кинетического подхода с использованием поворотно-изомерной модели для описания поведения полимерных цепей при наиболее широко исследованном квазистатическом механическом воздействии изложен ниже. С термодинамической точки зрения сила, приложенная к концам полимерной цепи в конформации статистического клубка при заданных термодинамических параметрах, в частности, в условиях равновесного изобарно-изотермического процесса, как функция деформации, или смещения концов цепи f(h) (см. Рис.1.4а) складывается из энергетического (энтальпийного) и энтропийного компонентов
где f(h) – сила как функция смещения h; ΔH – изменение энтальпии, ΔS – изменение энтропии, p – давление, T – температура. Изменение внутренней энергии или энтальпии полимерных цепей в изобарно-изотермических условиях связано с изменениями энергии, или потенциала взаимодействия первичных (ковалентных) и вторичных (преимущественно Ван-дер-ваальсовских) связей. Следовательно, энергетический (энтальпийный) компонент деформирующей силы, равен частной производной энергии, или потенциала соответствующей связи U по смещению (изменению ее длины), и при малом смещении относительно равновесного состояния пропорционален величине смещения:
где Δh(fU) – относительное изменение длины связи под действием силы fU, KU – коэффициент (модуль) энергетической упругости цепи. Такое поведение соответствует мгновенной равновесной энергетической упругости цепи, обусловленной деформированием ковалентных и физических связей и подчиняющейся закону Гука [ ]. Вследствие резкого различия в энергии (примерно на два десятичных порядка) ковалентных («жестких») и ван-дер-ваальсовских («мягких») связей значения KU для полимерных цепей могут различаться на несколько десятичных порядков в зависимости от того, какие связи деформируются, т.е. в зависимости от конформации цепи и направления приложения силы. С повышением температуры величина KU уменьшается пропорционально тепловому расширению, а при повышении давления – увеличивается пропорционально объемному сжатию. Так как характерное время развития таких деформаций пропорционально периоду высокочастотных тепловых колебаний (порядка 10-13-10-14 с), то очевидно, что энергетическая упругость полимерных цепей является практически мгновенной, т.е. при реальных длительностях механических воздействий имеет нереласационный характер. Очевидно, что энергетическая упругость полимерных цепей может проявляться только при замороженном сегментальном движении. При реализации такого движения основной вклад в упругое деформирование полимерных цепей вносит энтропийная упругость. В термодинамически равновесных условиях изобарно-изотермического процесса уменьшение энтропии цепи под воздействием силы fs определяется уменьшением числа возможных состояний при увеличении расстояния между концами цепи до
Сила упругого сопротивления цепи изменению ее конформации соответствует энтропийному компоненту общей силы сопротивления деформированию системы:
где Важнейшей особенностью энтропийной упругости полимерных цепей по сравнению с энергетической, помимо малых равновесных значений Кs и его возрастанию при повышении температуры, является низкая скорость процесса изменения конформаций цепи, обусловленная большим числом и размерами сегментов, совершающие крупномасштабные низкочастотные «транс»-«гош» переходы., от которой зависит Время, требуемое для перехода звеньев из одного состояния в другое, т.е.период или величина, обратная частоте колебаний сегментального движения, характеризующее кинетическую, или динамическую гибкость полимерных цепей в поворотно-изомерной модели, определяется величиной энергетического барьера ΔЕ, разделяющего «транс» и «гош» состояния:
При ΔЕ <<kT это время мало и с повышением высоты энергетического барьера оно экспоненциально возрастает. При этом проявляется значительное необратимое рассеяние (диссипация) энергии вследствие избыточного производства энтропии (внутреннего трения). Возникающая при сила внутреннего трения fξ пропорциональна скорости изменения расстояния между концами цепи
где Очевидно, что при большой длине цепи (большом N) основной вклад в коэффициент и, следовательно, в силу трения вносит компонент трения о вязкую среду, пропорциональный N. Возникающая при изменении конформации цепи со скоростью
Характерное время задержки деформации, или время релаксации процесса изменения конформаций цепи θ, определяется как отношение коэффициента внутреннего трения и энтропийной упругой константы цепи [Де Жен,с.201]:
Из уравнения (1.15) следует, что величина θ пропорциональна Квадратичная зависимость θ от длины цепи получена также в классической модели Рауза для растяжения идеально-гибкой цепи в вязкой среде [Тоб.]. В соответствии с этой моделью идеально-гибкая цепь, погруженная в среду с вязкостью ηs, при растяжении под действием силы fs разбивается на N участков (субцепей, или бусин, «блобов»), имеющих форму статистического клубка со средне-квадратичным размером
где Bp и θр – податливость и время задержки деформации р-й моды соответственно. Набор (спектр) времен задержки θр при этом определяется соотношением:
где ξ – коэффициент трения бусин о вязкую среду, равный в соответствие с законом Стокса:
Минимальное время задержки соответствует первой моде (р=1), а максимальное – конечной (р=N-1). Очевидно, что в отличие от поворотно-сегментальной модели в реальных гибких полимерных цепях наряду с индивидуальным сегментальным движением (β формой), возможна также кооперативная α форма, при которой возможно взаимодействие и корреляция сегментов друг с другом с возрастанием времени релаксации. При замороженной индивидуальной или кооперативной форме сегментального теплового движения определяющую роль должны играть мелкомасштабные формы внутримолекулярного теплового движения, причем каждой дополнительной форме сегментального движения должен соответствовать свой спектр времен релаксации, и среднее значение времени релаксации при прочих равных условиях должно уменьшаться от α к
Помимо высоко- и низкочастотных форм внутримолекулярного (микроброуновского) теплового движения и соответствующей им мгновенной и замедленной упругости полимерные цепи способны к различным формам макроброуновского теплового движения цепей как целого, в первую очередь поступательной (трансляционной) и вращательной самодиффузии, активируемой внешним воздействием. Такое движение играет определенную роль в процессах диэлектрической релаксации, но особенно важно оно в процессах механической релаксации, обусловливая протекание неравновесных необратимых процессов вязкого течения расплавов и растворов полимеров и полимерных композиций при медленных квазистатических или низкочастотных режимах. Если микроброуновское движение проявляется при любой молекулярной топологии полимеров (макромолекулярной линейной или разветвленной и сетчатой) и длине цепей, то макроброуновское движение возможно только в случае макромолекул конечных размеров - линейных и слабо разветвленных макромолекул конечной длины,не связанных друг с другом в непрерывную сетку прочными ковалентными или физическими связями. Скорость
Подстановка этого значения μt в диффузионное уравнение Эйнштейна
Вращательная подвижности макромолекулярного клубка как целого связана с его гидродинамическим радиусом Rg, величина которого пропорциональна Характерные времена трансляционной и вращательной диффузионной подвижности макромолекулярного клубка как целого, так называемые фундаментальные времена релаксации, равные времени, необходимом клубку для перемещения на расстояние, равное h или поворота какой-либо точки на поверхности клубка на расстояние, равное Rg, соответственно, определяются отношением Диффузионная подвижность невозмущенного макромолекулярного клубка как целого встречается очень редко, например, в разбавленных растворах полимеров при воздействии механических сил. Более важным и сложным типом макроброуновского движения цепей является поступательное (трансляционное) движение индивидуальной («меченой») цепной макромолекулы в лабиринте себе подобных, определяющее вязкое течение расплавов и концентрированных растворов полимеров. Одной из моделей такого движения является проползание гибкой цепи между фиксированными препятствиями (другими цепями), образующими некоторую трубку, в которой находится «меченая» цепь и длина которой равна контурной длине цепи L=Nls., где N и ls , соответственно, число сегментов и их длина (Рис.1.6).
(а) (б)
Рис.1.6. Схематическое изображение гибкой цепи в «трубке» (а) и элементарного акта рептации (б) [Де Жен, с.253].
Подвижность цепи в трубке характеризуется коэффициентом трансляционной диффузии цепи вдоль трубки Dtt, который, в соответствие с диффузионным соотношением Эйнштейна, связан с подвижностью сегмента μs и его коэффициентом диффузии De (величиной порядка
Характерным временем такого движения (рептации) служит макроскопическое время релаксации θm, равное времени, необходимому для выползания макромолекулы из первоначальной трубки:
где θе – продолжительность элементарного акта движения сегмента (величина порядка За время θм цепь перемещается вдоль трубки на расстояние, равное контурной длине цепи (Nls), тогда как в пространстве она перемещается на расстояние, равное средне-квадратичному расстоянию между ее концами (
При De= 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2024 stydopedia.ru Все материалы защищены законодательством РФ.
|
 с, что соответствует режиму ультразвуковых колебаний с частотой порядка
с, что соответствует режиму ультразвуковых колебаний с частотой порядка  Гц, и высокочастотные колебания, активируемые механической силой, обусловливают практически мгновенное (за время, близкое к периоду колебаний) развитие небольших обратимых (равновесных упругих) деформаций, пропорциональных действующей силе.
Гц, и высокочастотные колебания, активируемые механической силой, обусловливают практически мгновенное (за время, близкое к периоду колебаний) развитие небольших обратимых (равновесных упругих) деформаций, пропорциональных действующей силе. от числа звеньев в нейNпри их длине l0, аналогичную расстоянию, которое проходит молекула при свободном блуждании за N скачков при длине свободного пробега l0:
от числа звеньев в нейNпри их длине l0, аналогичную расстоянию, которое проходит молекула при свободном блуждании за N скачков при длине свободного пробега l0: (1.2)
(1.2) ) к статистически рассчитанному для идеально гибкой цепи:
) к статистически рассчитанному для идеально гибкой цепи: (1.3)
(1.3)
 - персистентная длина)
- персистентная длина) ), определяемая по соотношению:
), определяемая по соотношению: - длина цепи по контуру (L=Nl0), и степень полимеризации статистического сегмента NА=A/l0. Значения А и NА соответствуют эффективной длине участков реальной цепи, сочетание которых обусловливает ее поведение, описываемое статистикой идеальной свободно-сочлененной цепи.
- длина цепи по контуру (L=Nl0), и степень полимеризации статистического сегмента NА=A/l0. Значения А и NА соответствуют эффективной длине участков реальной цепи, сочетание которых обусловливает ее поведение, описываемое статистикой идеальной свободно-сочлененной цепи. связей главной цепи без изменения фиксированных углов. Эти повороты имеют активационный характер и связанны с переходами сегvентов между рядом дискретных поворотных изомеров (конформеров), обладающих минимумом энергии.
связей главной цепи без изменения фиксированных углов. Эти повороты имеют активационный характер и связанны с переходами сегvентов между рядом дискретных поворотных изомеров (конформеров), обладающих минимумом энергии. относительно их «транс»-положения (плоского зигзага) соответствует двум другим устойчивым («гош») конформерам сегмента. Разность уровней энергий «транс» и «гош»-конформеров Δе характеризует статическую (равновесную, или термодинамическую) жесткость цепи, а величина потенциального барьера ΔЕ – ее кинетическую жесткость. Величина этих параметров определяется количеством и типом σ-связей в цепи, наличием циклов, жестких кратных связей и их сопряжений, особенно в ароматических циклах, а также энергией вторичных физических связей.
относительно их «транс»-положения (плоского зигзага) соответствует двум другим устойчивым («гош») конформерам сегмента. Разность уровней энергий «транс» и «гош»-конформеров Δе характеризует статическую (равновесную, или термодинамическую) жесткость цепи, а величина потенциального барьера ΔЕ – ее кинетическую жесткость. Величина этих параметров определяется количеством и типом σ-связей в цепи, наличием циклов, жестких кратных связей и их сопряжений, особенно в ароматических циклах, а также энергией вторичных физических связей. , (1.5)
, (1.5) (1.6)
(1.6) (1.7)
(1.7)


 (см. рис.3, схема 2). Изломы образуются за счет переходов сегментов из «транс-» в «гош»-конформации («транс» - «гош» переходов). Доля «гош»-конформеров, или вероятность образования излома цепи служит универсальным критерием гибкости цепи (критерием Флори)
(см. рис.3, схема 2). Изломы образуются за счет переходов сегментов из «транс-» в «гош»-конформации («транс» - «гош» переходов). Доля «гош»-конформеров, или вероятность образования излома цепи служит универсальным критерием гибкости цепи (критерием Флори)  . В первом приближении
. В первом приближении  0 иуглом
0 иуглом  (1.8)
(1.8) и длиной статистического сегмента A) следующим соотношением:
и длиной статистического сегмента A) следующим соотношением: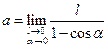 ; А=2а (1.9)
; А=2а (1.9) гибридных облаков) в цепи φар.
гибридных облаков) в цепи φар.
 - расстояние между концами цепи;
- расстояние между концами цепи;  - длина складки;
- длина складки;  (1.11)
(1.11) , (1.12)
, (1.12) и
и  соответственно, т.е. разности частной производной обратимого изменения внутренней энергии или энтальпии по смещению и произведения температуры на производную изменения энтропии по нему:
соответственно, т.е. разности частной производной обратимого изменения внутренней энергии или энтальпии по смещению и произведения температуры на производную изменения энтропии по нему: , (1.13)
, (1.13) , (1.14)
, (1.14) (fs) относительно равновесного расстояния h0 и в поворотно-изомерной модели цепи, состоящей из N гибких сегментов длиной А, описывается соотношением:
(fs) относительно равновесного расстояния h0 и в поворотно-изомерной модели цепи, состоящей из N гибких сегментов длиной А, описывается соотношением: (1.15)
(1.15) , (1.16)
, (1.16) - коэффициент (модуль) равновесной энтропийной упругости полимерной цепи, пропорциональный температуре и обратно пропорциональный длине полимерной цепи. По абсолютной величине Кs значительно меньше модуля энергетической упругости Кu.
- коэффициент (модуль) равновесной энтропийной упругости полимерной цепи, пропорциональный температуре и обратно пропорциональный длине полимерной цепи. По абсолютной величине Кs значительно меньше модуля энергетической упругости Кu. (1.10)
(1.10) , а коэффициент пропорциональности, или коэффициент молекулярного, или сегментального трения ξ, складывается из трех компонентов: первый обусловлен трением сегментов о вязкую окружающую среду и пропорционален числу сегментов, второй - преодолением потенциальных барьеров при «транс»-«гош» переходах и обратно пропорционален числу сегментов, и третий - трением друг о друга достаточно удаленных сегментов (трением Серфа) и не зависящий от числа сегментов [Де Жен,с.222]. Тогда суммарный (эффективный) коэффициент сегментального трения при заданной скорости изменении конформации цепи равен:
, а коэффициент пропорциональности, или коэффициент молекулярного, или сегментального трения ξ, складывается из трех компонентов: первый обусловлен трением сегментов о вязкую окружающую среду и пропорционален числу сегментов, второй - преодолением потенциальных барьеров при «транс»-«гош» переходах и обратно пропорционален числу сегментов, и третий - трением друг о друга достаточно удаленных сегментов (трением Серфа) и не зависящий от числа сегментов [Де Жен,с.222]. Тогда суммарный (эффективный) коэффициент сегментального трения при заданной скорости изменении конформации цепи равен: , (1.17)
, (1.17) - коэффициенты трения сегментов о вязкую среду, трения при преодолении потенциального барьера «транс»-«гош» перехода и трения Серфа, соответственно, приходящиеся на один сегмент.
- коэффициенты трения сегментов о вязкую среду, трения при преодолении потенциального барьера «транс»-«гош» перехода и трения Серфа, соответственно, приходящиеся на один сегмент. (1.18)
(1.18) (1.19)
(1.19) , т.к. ξ при определяющем вкладе компонента трения о вязкую среду пропорционален N, а Ks – обратно пропорционален N (см. уравнение (1.12), т.е. решающее влияние на время релаксации сегментальной подвижности цепи оказывает ее длина.
, т.к. ξ при определяющем вкладе компонента трения о вязкую среду пропорционален N, а Ks – обратно пропорционален N (см. уравнение (1.12), т.е. решающее влияние на время релаксации сегментальной подвижности цепи оказывает ее длина. и связанных упругими «мягкими» пружинами с модулем упругости Ks. Движение концов цепи (ожерелья бусин, соединенных пружинами) относительно ее центра тяжести, т.е. упругое смещение под действием силы fs, разбивается на формы (моды) р, число которыхравно N-1, с характерным временем задержки деформации для каждой моды, т.е. со спектром времен задержки для цепи как целого, состоящим из р компонентов. Общее смещение конца цепи (N-й бусины) во времени uN(t) описывается при этом дискретной функцией [Малк.]:
и связанных упругими «мягкими» пружинами с модулем упругости Ks. Движение концов цепи (ожерелья бусин, соединенных пружинами) относительно ее центра тяжести, т.е. упругое смещение под действием силы fs, разбивается на формы (моды) р, число которыхравно N-1, с характерным временем задержки деформации для каждой моды, т.е. со спектром времен задержки для цепи как целого, состоящим из р компонентов. Общее смещение конца цепи (N-й бусины) во времени uN(t) описывается при этом дискретной функцией [Малк.]: , (1.20)
, (1.20) , (1.21)
, (1.21) (1.22)
(1.22) форме сегментального движения. При этом времена релаксации процессов изменения конформаций цепей в результате внутримолекулярного теплового движения, резко зависящие от температуры, которая входит в уравнения (1.15 и 1.17 не только непосредственно, но и косвенно через вязкость среды или коэффициенты молекулярного трения. При больших N эти временаимеют макроскопические значения и соизмеримы с характерными временами механических воздействий в методах механической и диэлектрической релаксационной спектроскопии.
форме сегментального движения. При этом времена релаксации процессов изменения конформаций цепей в результате внутримолекулярного теплового движения, резко зависящие от температуры, которая входит в уравнения (1.15 и 1.17 не только непосредственно, но и косвенно через вязкость среды или коэффициенты молекулярного трения. При больших N эти временаимеют макроскопические значения и соизмеримы с характерными временами механических воздействий в методах механической и диэлектрической релаксационной спектроскопии.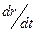 поступательного движения идеально-гибкого макромолекулярного клубка со средне-квадратичным расстоянием между концами
поступательного движения идеально-гибкого макромолекулярного клубка со средне-квадратичным расстоянием между концами  (1.23)
(1.23) дает следующее выражение для коэффициента трансляционной диффузии клубка Dt:
дает следующее выражение для коэффициента трансляционной диффузии клубка Dt: (1.24)
(1.24) и
и  к соответствующему кэффициенту диффузии Dt и Dr. Эти времена в решающей степени зависят от размеров клубка и вязкости среды и имеют макроскопические значения. Так, при вязкости среды 10 Па.с и h=50нм фундаментальное время релаксации клубка равно 0,2 с.
к соответствующему кэффициенту диффузии Dt и Dr. Эти времена в решающей степени зависят от размеров клубка и вязкости среды и имеют макроскопические значения. Так, при вязкости среды 10 Па.с и h=50нм фундаментальное время релаксации клубка равно 0,2 с.

 см2/с) следующим выражением:
см2/с) следующим выражением: (1.25)
(1.25) , (1.26)
, (1.26) с). При N=
с). При N=  время θм равно примерно 10 с.
время θм равно примерно 10 с.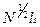 ). Следовательно, реальный коэффициент трансляционной диффузии Dt цепи по рептационному механизму равен:
). Следовательно, реальный коэффициент трансляционной диффузии Dt цепи по рептационному механизму равен: (1.27)
(1.27) см2/с и
см2/с и  Dt=
Dt=  cм2/с. При таком коэффициенте трансляционной диффузии цепь преодолеет за 10 суток расстояние
cм2/с. При таком коэффициенте трансляционной диффузии цепь преодолеет за 10 суток расстояние  =0,3 мм.
=0,3 мм.