|
|
Топоизомеразы и транскрипция геновПолагают, что репликация ДНК происходит на ядерном матриксе и один из связанных с матриксом ферментов обеспечивает раскручивание спирали ДНК. Нужен ли подобный фермент для транскрипции РНК? Недавние исследования показали, что раскручивание спирали ДНК играет важную роль, способствуя транскрипции РНК. Чтобы цепи ДНК разошлись, транскрипционно активный хроматин должен быть закручен (Ryoji, Worcel, 1984), и это закручивание осуществляется с помощью суперспирализации ДНК (рис. 12.32). Если суперспирализованные кольцевые плазмиды инъецировали в ядра ооцитов Xenopus, то эти плазмиды транскрибировались. Если же эти плазмиды линеаризовали (раскрывая их с помощью рестриктазы), то после инъекции в клетки они не транскрибировались (Harland et al., 1983; Pruitt, Reeder, 1984). Показано, что чувствительные к ДНКазе I сайты в активных генах возникают только в тех случаях, когда эти гены находятся под торзионным напряжением (Villeponteau et al., 1984). Ферментом, который ответствен за скручивание ДНК, позволяющее цепям ДНК разделиться, является топоизомераза II. С помощью антител к этому белку продемонстрировано, что топоизомераза II расположена в комплексе внутреннего ядерного матрикса с ядерной оболочкой (Berrios et al., 1985). Когда к ядерным препаратам добавляется ингибитор топоизомеразы II новобиоцин, напряжение с генов снимается, сайты, чувствительные к ДНКазе, исчезают, и транскрипция прекращается (Han et al., 1985; Glikin et al., 1984). Прикрепление ДНК к матриксу может быть необходимо также для предотвращения свободного вращения ДНК, что позволяет связанным с матриксом топоизомеразам скручивать хроматин (Cockerill, Garrard, 1986). Ζ-ДНК и петлевые структуры Одним из результатов суперспирализации может быть переход ДНК в альтернативную конформацию, отличающуюся от стандартной B-формы двойной спирали Уотсона–Крика. Суперспирализация
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 170_______________ ГЛАВА 12________________________________________________________________________
ДНК может привести к образованию «левозакрученной» Z-ДНК или петлевых структур. Образование петель постулируется в тех случаях, когда по термодинамическим причинам может происходить спаривание двух комплементарных участков одной цепи ДНК с большей вероятностью, чем спаривание двух цепей. На рис. 12.33 показано возможное спаривание оснований внутри одной цепи на 5'-конце гена коллагена 22 курицы. Высказано предположение (Larsen, Weintraub, 1982), что такие петлевые структуры являются сайтами узнавания для транс-регуляторных белков, и эти белки, будучи связанными, предотвращают образование нуклеосом в этой области ДНК. Другой конформационный переход, который может происходить под действием скручивания, это переход в Z-ДНК (Johnston, Rich, 1985; Kohwi-Shigematsu, Kohwi, 1985). Большая часть ядерной ДНК представлена правозакрученной двойной спиралью в B-форме, описанной Уотсоном и Криком. Одна-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ_____________________________ 171
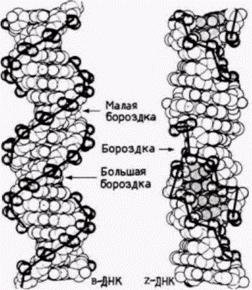
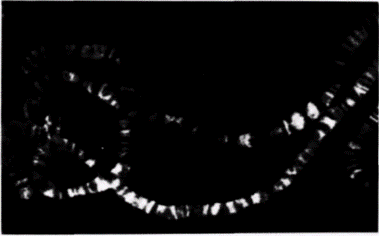
ко Рич и его коллеги обнаружили, что определенные ЦГ-богатые последовательности, такие как ЦГЦГЦГ, образуют предпочтительно левозакрученные двойные спирали (Wang et al., 1979). Физические свойства этой левозакрученной структуры отличаются от свойств стандартной ДНК. Левозакрученная спираль содержит 12 пар оснований на 1 виток (а не 10) и принимает конформацию «зиг-заг» (от· сюда название «Z-ДНК»). Помимо этого в Z-ДНК нет большой бороздки, которая обеспечивает присоединение ряда молекул к B-ДНК (рис. 12.34). Такая левозакрученная Z-ДНК обнаружена в геномах млекопитающих и дрозофилы. С помощью антител к Z-ДНК было показано, что Z-ДНК имеется на всем протяжении политенных хромосом дрозофилы (рис. 12.35). При этом она локализована в зонах между дисками. Считают, что эти междисковые зоны контролируют экспрессию генов, лежащих внутри дисков (Nordheim et al., 1981). Вначале полагали, что переход из B-ДНК в Z-ДНК может происходить только в условиях высокой концентрации солей. Однако позже было показано(Behe, Felsenfeld, 1981), что Z-ДНК формируется естественным образом при физиологических концентрациях солей, если остатки цитидинов в чередующихся ГЦ-полимерах метилированы. Таким образом, метилирование цитидина может позволить B-ДНК перейти в Z-конформацию. Кроме того, в отличие от В-ДНК, которая ускоряет формирование нуклеосом из гистонов. Z-ДНК эффективно подавляет формирование нуклеосом (Nickol et al., 1982). На самом деле гистоны будут связываться с Z-ДНК, но они не будут ассоциироваться в нуклеосомный октамер. РЕЗЮМЕ В данный момент у нас в руках части одной или нескольких головоломок. Мы не уверены, что имеем все части, и даже не знаем, сколько всего головоломок. Вполне вероятно, что существует несколько способов активации транскрипции. За прошедшие пять лет выяснилось, что образования «активных генов» еще недостаточно для транскрипции. Помимо этого требуется «активный хроматин». В настоящее время изучается взаимоотношение между альтернативными структурами генов (метилирован-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 172____________ ГЛАВА 12_______________________________________________________________________ ными и неметилированными; B-ДНК и петлями, или Z-ДНК) и альтернативными структурами хроматина (НMG-модифицированными нуклеосомами и немодифицированными; свободными от нуклеосом областями и областями, содержащими нуклеосомы). Участие гистона H1 в общем подавлении транскрипции и участие скручивания в активации специфических локальных участков представляются ключевыми элементами в регуляции генов. Остается выяснить, каким образом хроматин информируется о том, какую структуру ему следует принять. Вероятно, существуют определенные реципрокные взаимодействия между ДНК и хроматином. Посредством этих взаимодействий ДНК может управлять формообразованием хроматина в конкретном направлении, а данное изменение в структуре хроматина может в свою очередь позволить ДНК изменить свою структуру определенным образом. Одна из моделей заключается в следующем: когда ДНК реплицируется, нуклеосомы разрушаются. На этом этапе характер метилирования может быть изменен или сохранен 1. В активных генах некоторые из транс-регуляторных факторов (такие, как ТАТА-связывающий белок) могут присоединяться к ДНК и эти участки ДНК избегнут конденсации с участием гистона H1. Вместе с тем нуклеосомы в гипометилированных участках могут изменить конформацию так, что окажутся способными присоединить HMG 14 и HMG 17. Экспонированная ДНК может затем связаться с топоизомеразами на ядерном матриксе. Это присоединение позволит ДНК закрутиться таким образом, что в участках промоторных и энхансерных последовательностей возникнут альтернативные структуры. В этих местах цис-последовательности могли бы затем присоединить транс-регуляторные белки и РНК-полимеразу для осуществления транскрипции. Это лишь одна из возможных моделей, удовлетворяющих экспериментальным данным, но и против нее имеются возражения. Как заметил Альбер Клод, мы только начали составлять опись обретенного нами богатства.
1 Вновь образующиеся нуклеосомы могут располагаться в соответствии с распределением метилированных оснований. ЛИТЕРАТУРА
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ______________________________ 173
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 174_______________ ГЛАВА 12_____________________________________________________________________________
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ 175
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 176_______________ ГЛАВА 12______________________________________________________________________________
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2024 stydopedia.ru Все материалы защищены законодательством РФ.
|