|
|
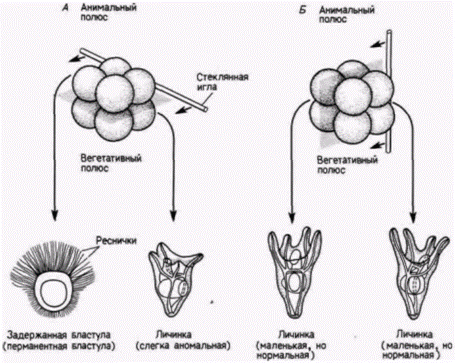
Свен Гёрстадиус: потенции и градиенты в ооцитеНо и Дриш оказался прав не на 100%. Как мы видели в предыдущей главе, существует много животных, развитие которых представляет собой главным образом мозаику самодифференцирующихся частей. Более важно, однако, что даже зародыши морского ежа не представляют собой коллекцию полностью эквипотенциальных клеток. Шведский биолог Свен Гёрстадиус в серии опытов, проведенных с 1928 по 1935 г.. отделял тонкими стеклянными иглами разные слои ранних зародышей морского ежа и наблюдал их последующее развитие (Hörstadius, 1928, 1939). Если он разделял 8-клеточный зародыш пополам по меридиану, проходящему через анимальный и вегетативный полюсы, то из обеих половин развивались личинки плутеусы, как и предсказывал Дриш. Но если зародыши на той же стадии разделяли пополам по экватору (т.е. на анимальную и вегетативную половины), то ни одна из частей не развивалась в нормальную личинку (рис. 8.5). Анимальная половина превращалась в полый шар, образованный реснитчатыми
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.
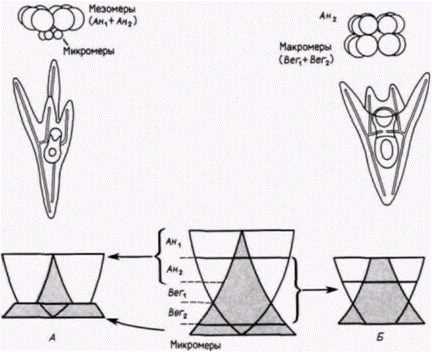
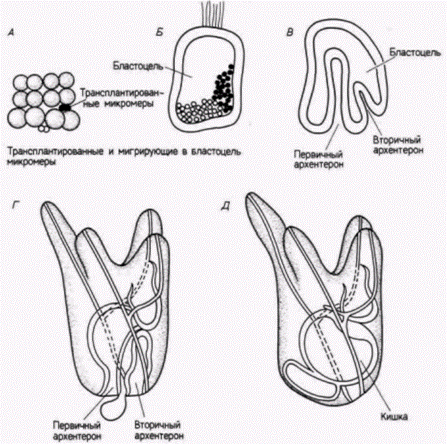
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ПРОГРЕССИВНАЯ ДЕТЕРМИНАЦИЯ_____________________________________________________ 45 эпидермальными клетками (называемый Dauerblastula – продолженная или задержанная бластула), а вегетативная половина развивалась в слегка аномальный зародыш с расширенной кишкой. Гёрстадиус смог повторить эти результаты, разрезая пополам яйца морского ежа и оплодотворяя обе половины по отдельности. У морских ежей фрагменты яиц (мерогоны) могут делиться и развиваться. даже если они имеют гаплоидное ядро. Если спермий проникает в половину, не содержащую гаплоидного ядра яйца, то мерогон также будет развиваться (рис. 8.6). Ко|да яйцо разрезали меридионально, из обеих половин яйца развивались нормальные зародыши. Однако когда яйцо разрезали экваториально, то после оплодотворения обеих половин из фрагментов формировались или окаймленный ресничками анимальный шарик, или зародыш с расширенной вегетативной кишкой. Следовательно, даже зародышам морского ежа, по-видимому, в некоторой степени присущ мозаицизм, по меньшей мере по анимально-вегетативной оси. Эти наблюдения побудили Гёрстадиуса осуществить, пожалуй, один из наиболее поразительных опытов в истории эмбриологии. Сначала (1935) он проследил нормальное развитие шести слоев клеток у 64-клеточного зародыша морского ежа. Как показано на рис. 8.7. А. анимальные клетки и первый вегетативный слой (Вег1) в норме образуют эктодерму, второй вегетативный слой (Вег2) дает начало энтодерме и части личиночной мезодермы, а из микромеров развивается мезодермальный скелет. Затем Гёрстадиус удалял у 64-клеточного зародыша оболочку оплодотворения, отделяя слои один от другого тонкими стеклянными иглами, и рекомбинировал их в разных сочетаниях. Изолированное анимальное полушарие превращалось в задержанную бластулу из эктодермальных клеток, покрытых ресничками (рис. 8.7. Б). Такой зародыш был назван анимализированным. Если Гёрстадиус комбинировал анимальное полушарие со слоем Вег1 (рис. 8.7. В), то получившаяся в результате личинка была менее анимализированной: у нее было подавлено образование ресничек и формировалась часть кишки. Если же анимальное полушарие было соединено со слоем Вег2 (рис. 8.7. Г), то развивалась личинка, выглядевшая нормальной. В этой комбинации слой Вег2, который в норме формирует только архентерон и его производные, теперь формировал также скелетные структуры. Сходным образом, когда анимальную половину комбинировали только с микромерами (рис. 8.7. Д), формировался маленький, выглядевший нормальным плутеус, но в этом случае энтодерма была полностью образована анимальными клетками. У такого плутеуса кишка возникала из клеток, которые в норме должны были образовать снабженную ресничками эктодерму. Эти опыты показали, что даже на 64-клсточной стадии анимальные клетки сохраняют генетический потенциал к образованию клеток кишки. Еще более важным было то, что способность к подавлению «анимализации» носит градиентный характер. Микромеры были более сильными «вегетализаторами», чем слой Вег2: но слой Вег2 в свою очередь был сильнее, чем слой Вег1. Следующая серия опытов Гёрстадиуса подтвердила существование анимализирующего фактора, который также распределен по градиенту. Микромеры комбинировали с каждым слоем 64-клеточного зародыша по отдельности. Изолированный слой Ан1(рис. 8.8. А) должен был образовывать задержанную бластулу. Когда к этому слою добавляли различное число изолированных микромеров, причем число их постоянно увеличивали, развивались все более полные зародыши. Комбинация только слоя Ан1. с четырьмя микромерами приводила к образованию нормального плутеуса. Из слоя Ан2, нормальный плутеус развивался уже при комбинации с двумя микромерами (рис. 8.8. Б), а добавление четырех микромеров приводило к аномальному увеличению энтодермальных структур. Из слоя Вег1без добавления микромеров образуется также только задержанная бластула. Однако добавление даже одного микромера вызывало резкое увеличение кишки (рис. 8.8. В), а изолированный слой Вег2вообще имел тенденцию к вегетализации без добавления каких-либо микромеров (рис. 8.8. Г). Таким образом, существует, по-видимому, градиент анимализации, усиливающийся по направлению от Ан1, к микромерам. Наиболее логичным объяснением этих результатов было предположение, что существуют два противоположно направленных градиента: вегетализирующий градиент с максимумом активности на вегетативном полюсе и анимализирующий градиент с максимальной активностью на анимальном полюсе. Именно такую систему двойных градиентов предложил старший коллега Гёрстадиуса Дж. Рунстрем (Runnstrom. 1929). Поскольку для развития более важны относительные, а не абсолютные концентрации различных веществ, рекомбинация бластомеров из двух полюсов восстанавливает все промежуточные значения градиентов. Сходным образом промежуточные клетки, еще содержащие максимум и минимум для обоих градиентов, также могут дать начало нормальному плутеусу (рис. 8.9). Эта модель двойного градиента оказалась очень полезной для объяснения других рекомбинационных опытов. Например, если микромеры из вегетативного полюса пересаживали в область, близкую к центру 32-клеточного зародыша, то они инвагинировали в бластоцель хозяина, где вызывали формирование хорошо развитого вторичного архентерона
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 46________________ ГЛАВА 8______________________________________________________________________________
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ПРОГРЕССИВНАЯ ДЕТЕРМИНАЦИЯ_____________________________________________________ 47
(рис. 8.10). Если же микромеры имплантировали на анимальный полюс, то формировалась очень маленькая вторичная кишка. Вместе с тем из зародышей не удалось выделить ни вегетативный, ни анимальный факторы. Ингибиторы белков (тяжелые металлы, NaSCN, краситель синий Эванса). по-видимому, уменьшали вегетализирующий градиент и таким способом анимализировали зародыш. Ингибиторы дыхания (CO, KCN, NaN3, Li) вегетализировали зародыш. Так, Убишу (Ubisch. 1929) удалось получить нормальный плутеус из анимальной половины, которая развивалась в растворе хлористого лития. Литий, по-видимому, ослабляет анимальную часть градиента в пределах анимальных клеток и тем самым способствует сбалансированному развитию этого полушария. Таким образом, мы имеем модель регуляционного развития, основанную на относительных концентрационных градиентах в ооците. Когда анимальные клетки комбинируют только с микромерами, формируется нормальный плутеус с клетками кишки, происходящими из анимальных клеток, которые в норме должны были формировать эктодерму. Следовательно, и на более поздних стадиях развития потенции клеток все еще шире, чем их судьба. Но из одних анимальных клеток теперь образуется только анимализированная бластула, так что способность одной клетки дать начало целой личинке исчезла. Другими словами, ее потенции стали ограниченными. Любая клетка с определенным соотношением анимальных и вегетативных веществ будет продуцировать определенный тип клеток. Следовательно, клетки анимального полюса с более высоким отношением анимального фактора к вегетативному в норме будут образовывать эктодермальную ткань. Однако некоторые ани-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ПРОГРЕССИВНАЯ ДЕТЕРМИНАЦИЯ______________________________________________________ 49 мальные клетки, рекомбинированные с сильным источником вегетативного фактора, будут формировать ткань кишки. Если зародыш разделен таким образом, что каждая половина содержит полный анимальный и вегетативный градиенты (т.е. разделение произведено по меридиану, проходящему вдоль анимально-вегетативной оси), то формируется полная личинка. Бластомеры остаются способными к регуляции до тех пор, пока не будут формироваться исключительно из анимальной или вегетативной цитоплазмы. Поэтому неудивителен тот факт, что после 32-клсточной стадии большая часть бластомеров по отдельности более не может дать начало целой личинке (Morgan. 1895), а микромеры с 16-клеточной стадии уже неспособны к этому (Hagstrom. Lonning, 1965: Okazaki. 1975). Даже для зародыша, развитие которого является регуляционным, наступает время, когда потенции его клеток становятся ограниченными. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2024 stydopedia.ru Все материалы защищены законодательством РФ.
|