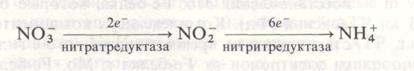|
|
Усвоение минеральных веществРастения для своего развития нуждаются в значительных количествах азота. Формы азота в окружающей растение среде чрезвычайно разнообразны: в атмосфере - молекулярный азот и пары аммиака, в почве - неорганические (нитраты, нитриты, аммоний, аммиак) и органические (аминокислоты, амиды, полипептиды и др.) соединения азота. Наиболее полно вопрос об использовании растениями нитратного и аммиачного азота разработан академиком Д.Н. Прянишниковым (1955) и его сотрудниками. Ими была установлена равноценность этих источников азота и изучены условия эффективного использования его высшими растениями. Одним из важнейших факторов, определяющих поглощение растениями неорганических форм азота, является реакция питательной среды. В слабокислой среде, при рН 5, лучше поглощаются нитраты. Наоборот, в нейтральной среде, при рН 7, преимущество имеет аммоний. Для усвоения аммония большое значение имеет и наличие в среде достаточного количества ионов кальция. Физиологическая особенность процессов усвоения заключается в том, что аммоний сразу после поглощения метаболизируется в корнях, превращаясь в азот аминокислот и амидов. Концентрация аммония в тканях и пасоке обычно очень низкая. Поэтому аммонийная форма азота эффективна только при условии высокой фотосинтетической активности или достаточного количества запасных углеводов. При недостатке органических кислот аммиак не успевает связываться и может быть токсичным для растения. Судьба поглощенного растениями нитрата может быть различной. Поступившие нитраты либо запасаются в вакуолях клеток корня, либо подаются с пасокой в надземную систему. Включаться в органические соединения они могут как в корнях, так и в листьях. Причем метаболизация нитратов начинается с их восстановления до аммония. На поглощение растениями нитратов и аммония существенно влияет температура среды. Причем реакция самых разных растений на температурные воздействия оказывается очень сходной: при пониженной температуре относительно или даже абсолютно больше поглощается аммония, чем нитрата, увеличивается метаболическая нагрузка корней по ассимиляции азота. Наиболее неустойчивым к низкой температуре этапом азотного обмена является загрузка ксилемы азотистыми соединениями, особенно нитратами. Замедление оттока приводит к снижению их поглощения. Таким образом, предпочтительность использования растениями разных форм минерального азота зависит от ряда условий (рН и ионного состава среды, наличия достаточного пула органических кислот, способных связывать аммиак, температуры и др.). Все эти факторы необходимо учитывать при разработке мероприятий по повышению эффективности применения азотных удобрений. Редукция нитрата. Поскольку в органические соединения включается только аммонийный азот, ионы нитрата, поглощенные растением, должны восстанавливаться в клетках до аммиака. Установлено, что процесс редукции нитрата в растениях осуществляется в два этапа: 1. Восстановление нитрата до нитрита (NO3- до NO2-), сопряженное с переносом двух электронов и катализируемое ферментом нитратредуктазой. 2. Восстановление нитрита до аммиака (NO2- до NH4+), сопряженное с переносом шести электронов и катализируемое ферментом нитритредуктазой. Суммарно процесс может быть изображен следующим образом:
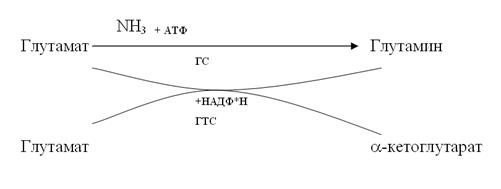
Нитратредуктаза представляет собой гем- и молибденсодержащий флавопротеин, участвующий в переносе электрона от НАД Н к NO3-. Восстановление нитратов до нитритов происходит в цитозоле клеток корня и листа. Не исключается возможность локализации нитратредуктазы на плазмалемме и на мембранах органелл, граничащих с растворимой фазой клетки. Но эти ассоциации фермента с мембранами удерживаются слабыми связями и легко разрушаются при выделении органелл. Вторая стадия восстановления минерального азота осуществляется при участии фермента нитритредуктазы. Это относительно низкомолекулярный белок, включающий около 600 аминокислотных остатков, который содержит железопорфириновую простетическую группу и железо в виде кластера 4Fe4S. В листьях нитритредуктаза локализована в хлоропластах и в качестве донора электронов использует восстановленный в световой фазе фотосинтеза ферридоксин (Фд). В корнях NO2- восстанавливается в пропластидах с использованием НАДФ-Н, образующегося в пентозофосфатном пути дыхания. Восстановление нитратов у растений может осуществляться и в листьях, и в корнях, однако относительная доля участия этих органов в редукции нитратов у растений разных видов сильно варьирует. По этому признаку растения подразделяют на три основные группы: 1. Растения, практически полностью восстанавливающие нитраты в корнях и транспортирующие азот к листьям в органической форме. К этой группе относятся многие древесные растения, а также некоторые представители сем. Ericaceae и Vacciniaceae (черника, клюква), многие виды Rhododendron. 2. Растения, практически не проявляющие нитратредуктазной активности в корнях и ассимилирующие нитраты в листьях. Это дурнишник (Xanthium), некоторые виды Borago. К этой группе примыкают хлопчатник и представители сем. Chenopodiaceae (свекла, марь), у которых основное количество поглощенного нитрата восстанавливается в листьях. 3. Растения, способные поддерживать активность нитратредуктазы и в листьях, и в корнях. Ассимиляция нитратов в листьях на свету тесно связана с процессом фотосинтеза. Реакции фотосинтеза используются как источник АТР для синтеза нитрат- и нитритредуктазы и транспорта нитратов, а также как источник восстановителей (для функционирования этих ферментов) и субстрата (органических кислот) для связывания конечного продукта восстановления - аммиака. Ассимиляция аммиака. Аммоний, поступив в корни растения, здесь же довольно быстро с помощью реакций аминирования (присоединения к органическим кислотам) используется на синтез ряда аминокислот, основных «кирпичиков» для синтеза белков. Так, из пировиноградной кислоты образуется аланин, из щавелевоуксусной и фумаровой - аспарагиновая кислота, из α- кетоглутаровой - глутаминовая кислота. В этих реакциях особенно наглядно проявляется тесная связь между минеральным питанием растений и дыханием, промежуточными продуктами которого являются названные органические кислоты. При обильном поступлении иона аммония в корнях происходит процесс амидирования аспарагиновой и глутаминовой аминокислот с образованием соответственно аспарагина и глутамина. Основной путь первичной ассимиляции аммония, включающий две последовательные сопряженные реакции, катализируемые ферментами глутаминсинтетазой (ГС) и глутаматсинтетазой (ГТС).
Механизм реакции включает перенос амидной группы глутамина на α-кетоглутарат в присутствии восстановителя, в результате чего происходит образование двух молекул глутамата. Глутаминсинтетаза (ГС) катализирует реакцию, в которой глутамат функционирует как акцептор NН3 для образования глутамина. Для этой реакции требуется АТФ. Причем ГС обладает гораздо большим сродством к NН3, чем глутаматдегидрогеназа (ГДГ). В настоящее время ГС - ГТС-путь считается главным в ассимиляции аммиака, образующегося в хлоропласте в результате восстановления нитратов. ГДГ имеет преимущества в темноте и в условиях аммонийного питания растений. Функционируют обе системы как в листьях, так и в корнях. Более простым способом связывания аммиака является образование аммонийных солей органических кислот. У растений с кислым клеточным соком (щавель, бегония, осоки, хвощи) содержание аммонийного азота в десятки раз превосходит содержание амидного азота. Аммиак в растениях обезвреживается и при образовании мочевины. Исходным продуктом для синтеза мочевины служит аминокислота орнитин, которая, присоединяя аммиак и углекислоту, превращается в цитруллин. Цитруллин присоединяет еще одну молекулу аммиака и образует аргинин. Аргинин под действием фермента аргиназы с участием воды расщепляется на орнитин и мочевину. Мочевина неядовита для растений, хорошо усваивается, использование ее азота для всевозможных синтетических процессов происходит очень легко, так как в растительных тканях имеется фермент уреаза, катализирующий расщепление мочевины. Усвоение фосфора и серы. В растительных тканях фосфор присутствует в органической форме и в виде ортофосфорной кислоты и ее солей. Он входит в состав белков (фосфопротеинов), нуклеиновых кислот, фосфолипидов, фосфорных эфиров сахаров, нуклеотидов, принимающих участие в энергетическом обмене (АТР, NAD+ и др.), витаминов и многих других соединений. Основной запасной формой фосфора у растений является фитин - кальций-магниевая соль инозитфосфорной кислоты (инозитолгексафосфата): Значительные количества фитина (0,5 - 2% на сухую массу) накапливаются в семенах, составляя до 50% от общего фосфора в них. Радиальное передвижение фосфора в зоне поглощения корня до ксилемы происходит по симпласту, причем его концентрация в клетках корня в десятки - сотни раз превышает концентрацию фосфата в почвенном растворе. Транспорт по ксилеме осуществляется в основном или полностью в форме неорганического фосфата; в этом виде он достигает листьев и зон роста. Фосфор, как и азот, легко перераспределяется между органами. Из клеток листьев он поступает в ситовидные трубки и по флоэме транспортируется в другие части растения, особенно в конусы нарастания и в развивающиеся плоды. Аналогичный отток фосфора происходит и из стареющих листьев. Сера, как и все биогенные элементы, участвует в биологическом круговороте веществ. Автотрофные растения поглощают серу в виде высшего окисла SО42-, восстанавливая его до уровня SH-групп органических веществ. В корнях или в листьях сульфаты очень быстро восстанавливаются до сульфидов по схеме: SO42- → SO32- → S2- Процесс восстановления сопровождается изменением валентности атома серы. Химизм процесса во всех деталях у растений изучен еще недостаточно. Более подробно известно о первом этапе, восстановлении сульфата в сульфиты. Сульфат с помощью молекул АТФ активируется с образованием аденозин-5фосфосульфата (АФС) и пирофосфорной кислоты. После этого АФС восстанавливается до сульфита и аденозинмонофосфата. С помощью метода меченых атомов удалось показать, что минеральная сера уже через несколько минут может входить в состав аминокислот цистеина и метионина. При распаде белков сера снова окисляется до сульфатов и в таком виде передвигается к местам, где возникает острая потребность в сере. Синтетическая деятельность корня. Корни являются органами: · поглощения воды и минеральных элементов; · синтеза сложных соединений. В ряде научных работ (Д.А. Сабинин, А.А. Шмук, А.Л. Курсанов, К. Мотес и др.) показано, что в корневых системах растений синтезируется да 16 аминокислот, белки, нуклеиновые кислоты и их производные, предшественники пластидных элементов, ряд витаминов и физиологически активных веществ, каучук и др. В пасоке бобовых растений весь поглощенный корнями азот находится в органической форме. В трахеидном соке таких древних по происхождению растений, как наши хвойные деревья, в частности сосны, обнаружена 6 аминакислот: аланин, аспарагиновая, треонин, глицин, валин, серин. При этом следует отметить, что неблагоприятные условия, например корневая аноксия, снижают синтез в корнях сосны ряда указанных аминокислот. В корнях растений синтезируется и ряд весьма специфических веществ. Так, в корневых системах растений табака синтезируется алкалоид никотин азотсодержащее гетероциклическое соединение. Образовавшись в корнях, как и большинство алкалоидов, никотин вместе с восходящим током передвигается в надземную часть и накапливается в листьях. Здесь содержание его достигает 1 – 10% массы сухого вещества. Корни растений являются местам синтеза фитогормонов, в частности цитокининов, без которых невозможен рост стебля. Энергично растущие ветки, если их срезать и поместить в питательный раствор, сразу же прекращают рост, который не возобновляется да тех пор, пока не появятся придаточные корни. При старении листьев часть органических веществ, в том числе белков, распадается, а продукты распада направляются в стебли и корни. Образующийся при этом аммиак обезвреживается, включаясь в состав амидов и новых аминакислот. К сказанному необходимо добавить, что корням свойственна и функция выделения веществ. Основным корневым выделением является углекислота, образующаяся в процессе дыхания корней. Реагируя с водой, СО2 дает очень слабую угольную кислоту, которая, однако, способна подкислять среду и тем самым переводить часть недоступных минеральных веществ почвы в доступные для растений формы. Осуществляя многочисленные синтезы и превращения веществ, корни растений способны не только их накапливать, но и частично выделять в почву. Наряду с углекислотой могут выделяться и некоторые органические кислоты, особенно яблочная, лиманная, винная, отдельные аминокислоты и некоторые другие соединения. Выделяется и небольшая часть минеральных элементов, в частности калия, кальция и фосфора. Общее количество корневых выделений мажет достигать 5 % всей массы синтезированных в растении веществ. Корневые системы древесных растений способны выделять да нескольких десятков различных веществ, что нельзя не учитывать при выяснении взаимного влияния древесных растений друг на друга в лесу и на микроорганизмы почвы. Как видим, деятельность корневых систем у растений является очень многосторонней. Поэтому на улучшение работы корней направлены, по существу, почти все приемы агротехники в сельском хозяйстве, многие – в лесном. Сюда относятся и правильная обработка почвы, постоянное ее рыхление, обеспечивающее аэрацию корневых систем, и сохранение в почве влаги, и орошение, и применение удобрений для оптимизации водного и минерального питания растений, и дренаж избыточно увлажненной почвы с целью улучшения снабжения корней свободным кислородом и т.д. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2024 stydopedia.ru Все материалы защищены законодательством РФ.
|