
|
|
Непрямой (вторичный) остеогистогенез. Развитие кости на месте хряща.На 2-м месяце эмбрионального развития в местах будущих трубчатых костей закладывается из мезенхимы хрящевой зачаток, который очень быстро принимает форму будущей кости (хрящевая модель). Зачаток состоит из эмбрионального гиалинового хряща, покрытого надхрящницей. Некоторое время он растет как за счет клеток, образующихся со стороны надхрящницы, так и за счет размножения клеток во внутренних участках. Развитие кости на месте хряща, т.е. непрямой остеогенез, начинается в области диафиза (т.н. перихондральное окостенение). Образованию перихондральной костной манжетки предшествует разрастание кровеносных сосудов. Происходит дифференцировка остеобластов, образующих в виде манжетки сначала ретикулофиброзную костную ткань (первичный центр окостенения), затем заменяющуюся на пластинчатую. Образование костной манжетки нарушает питание хряща. Вследствие этого в центре диафизарной части хрящевого зачатка возникают дистрофические изменения.Хондроциты вакуолизируются, их ядра пикнотизируются, образуются так называемые пузырчатые хондроциты. Рост хряща в этом месте прекращается. Удлинение перихондральной костной манжетки сопровождается расширением зоны деструкции хряща и появлением остеокластов, которые очищают пути для врастающих в модель трубчатой кости кровеносных сосудов и остеобластов. Это приводит к появлению очагов «внутреннего» эндохондрального окостенения(вторичные центры окостенения). В связи с продолжающимся ростом соседних неизмененных дистальных отделов диафиза хондроциты на границе эпифиза и диафиза собираются в продольные колонки. Таким образом, в колонке хондроцитов имеются два противоположно направленных процесса — размножение и рост в дистальных отделах диафиза и дистрофические процессы в его проксимальном отделе. Одновременно между набухшими клетками происходит отложение минеральных солей, обусловливающее появление резкой базофилии и хрупкости хряща. С момента разрастания сосудистой сети и появления остеобластов надхрящница перестраивается, превращаясь в надкостницу. В дальнейшем кровеносные сосуды с окружающей их мезенхимой, остеогенными клетками и остеокластами врастают через отверстия костной манжетки и входят в соприкосновение с обызвествленным хрящом. Под влиянием ферментов, выделяемых остеокластами, происходит растворение (хондролиз) обызвествленного межклеточного вещества. Диафизарный хрящ разрушается, в нем возникают удлиненные пространства, в которых «поселяются» остеоциты, образующие на поверхности оставшихся участков обызвествленного хряща костную ткань. Процесс отложения кости внутри хрящевого зачатка получил название эндохондрального, или энхондрального, окостенения (греч. endon — внутри). Одновременно с процессом развития энхондральной кости появляются и признаки ее разрушения остеокластами. Вследствие разрушения энхондральной костной ткани образуются еще большие полости и пространства (полости резорбции) и, наконец, возникает костномозговая полость. Из проникшей сюда мезенхимы образуется строма костного мозга, в которой поселяются стволовые клетки крови и соединительной ткани. В это же время по периферии диафиза со стороны надкостницы нарастают все новые и новые перекладины костной ткани, образующейся из надкостницы. Разрастаясь в длину по направлению к эпифизам и увеличиваясь в толщину, они образуют плотный (компактный) слой кости. Дальнейшая организация периостальной кости протекает иначе, чем организация энхондральной костной ткани. Вокруг сосудов, которые идут по длинной оси зачатка кости из прилегающей к ним мезенхимы, на месте разрушающейся ретикулофиброзной кости начинают образовываться концентрические пластинки, состоящие из параллельно ориентированных тонких коллагеновых волокон и цементирующего межклеточного вещества. Так возникают первичные остеоны. Просвет их широк, границы пластинок нерезко контурированы. Вслед за появлением первой генерации остеонов со стороны периоста начинается развитие общих (генеральных) пластинок, окружающих кость в области диафиза. Вслед за диафизом центры окостенения появляются в эпифизах. Этому предшествуют сначала дифференцировка хондроцитов, их гипертрофия, сменяемая ухудшением питания, дистрофией и кальцинацией. В дальнейшем отмечается процесс окостенения, подобный описанному выше. Оссификация сопровождается врастанием в эпифизы сосудов. В промежуточной области между диафизом и эпифизами сохраняется хрящевая ткань — метафизарный хрящ, являющийся зоной роста костей в длину. Рост трубчатых костей. Рост костей — процесс очень длительный. Он начинается у человека с ранних эмбриональных стадий и кончается в среднем к 20-летнему возрасту. В течение всего периода роста кость увеличивается как в длину, так и в ширину. Рост трубчатой кости в длину обеспечивается наличием метаэпифизарной хрящевой пластинки, в которой проявляются два противоположных гистогенетических процесса. Один — это разрушение эпифизарной пластинки с образованием костной ткани, а другой процесс — непрестанное пополнение хрящевой ткани путем новообразования клеток. Однако со временем процессы разрушения хрящевой ткани начинают преобладать над процессами новообразования, вследствие чего хрящевая пластинка истончается и исчезает. В метаэпифизарном хряще различают три зоны: · пограничную зону (интактного хряща), · зону столбчатых (активно делящихся) клеток и · зону пузырчатых (дистрофически измененных) клеток. Пограничная зона, расположенная вблизи эпифиза, состоит из округлых и овальных клеток и единичных изогенных групп, которые обеспечивают связь хрящевой пластинки с костью эпифиза. В полостях между костью и хрящом находятся кровеносные капилляры, обеспечивающие питанием клетки глубжележащих зон хрящевой пластинки. Зона столбчатых клеток содержит активно размножающиеся клетки, которые формируют колонки, расположенные по оси кости, и обеспечивают ее рост и длину. Проксимальные концы колонок состоят из созревающих, дифференцирующихся хрящевых клеток. Они богаты гликогеном и щелочной фосфатазой. Обе эти зоны наиболее реактивны при действии гормонов и других факторов, оказывающих влияние на процессы окостенения и роста костей. Зона пузырчатых клеток характеризуется гидратацией и разрушением хондроцитов с последующим эндохондральным окостенением. Дистальный отдел этой зоны граничит с диафизом, откуда в нее проникают остеогенные клетки и кровеносные капилляры. Продольно ориентированные колонки энхондральной кости являются по существу костными трубочками, на месте которых формируются остеоны. Впоследствии центры окостенения в диафизе и эпифизе сливаются и рост кости в длину заканчивается. Рост трубчатой кости в ширину осуществляется за счет периоста. Со стороны периоста очень рано начинает образовываться концентрическими слоями тонковолокнистая кость. Этот аппозиционный рост продолжается до окончания формирования кости. Количество остеонов непосредственно после рождения невелико, но уже к 25 годам в длинных костях конечностей количество их значительно увеличивается. 33. Мышечное волокно. Световая, поляризационная и электронная микроскопия. Миосателлитоциты. Основной структурно-функциональной единицей скелетных мышц является мышечное волокно. Это очень большое вытянутое многоядерное образование длиной в несколько сантиметров, при поперечнике около 100 мкм. Размер мышечного волокна в десятки тысяч раз превышает размер средней по величине клетки под микроскопом на продольном срезе мышечного волокна видна поперечная исчерченность, которая обусловлена тем, что его внутренние структуры периодически (через каждые 2—2,5 мкм) многократно повторяются. Подобные гигантские поперечно-полосатые волокна составляют мышечную ткань не только скелетной мускулатуры, но также сердца и некоторых внутренних органов. Миосателлитоциты - это малодифференцированные клетки, являющиеся источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются. Миосателлитоциты одноядерны, их ядра овальной формы и мельче, чем в симпластах. Они обладают всеми органеллами общего значения (в том числе и клеточным центром). Ядра миосимпластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами.
34. Миофибрилла. Саркомер, строение, формула саркомера. Молекулярная организация актиновых и миозиновых миофиламентов. Саркотубулярная система. Механизм мышечного сокращения. Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной. Длина всего волокна может измеряться сантиметрами при толщине всего 50—100 мкм. Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой. Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. У полюсов ядер располагаются органеллы общего значения — аппарат Гольджи и небольшие фрагменты гранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно. Саркомер — это структурная единица миофибриллы. Каждая миофибрилла имеет поперечные темные и светлые диски, имеющие неодинаковое лучепреломление (анизотропные A-диски и изотропные I-диски). Каждая миофибрилла окружена продольно расположенными и анастомозирующими между собой петлями агранулярной эндоплазматической сети — саркоплазматической сети, или саркоплазматического ретикулума. Соседние саркомеры имеют общую пограничную структуру — Z-линию (или телофрагму). Она построена в виде сети из белковых фибриллярных молекул, среди которых существенную роль играет альфа-актинин. С этой сетью связаны концы тонких, актиновых, филаментов. От соседних Z-линий актиновые филаменты направляются к центру саркомера, но не доходят до его середины. Филаменты актина объединены с Z-линией и нитями миозина фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомезина. Она образует в сечении М-линию, или мезофрагму. В узлах этой М-линии закреплены концы толстых, миозиновых филаментов. Другие их концы направляются в сторону Z-линий и располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина.
Альфа-актининовые сети Z-линий соседних миофибрилл связаны друг с другом промежуточными филаментами. Они подходят к внутренней поверхности плазмолеммы и закрепляются в кортикальном слое цитоплазмы, так что саркомеры всех миофибрилл располагаются на одном уровне. Это и создает при наблюдении в микроскоп впечатление поперечной исчерченности всего волокна. Источником ионов кальция служат цистерны агранулярной эндоплазматической сети. Они вытянуты вдоль миофибрилл около каждого саркомера и образуют саркоплазматическую сеть. Именно в ней аккумулируются ионы кальция, когда миосимпласт находится в расслабленном состоянии. На уровне Z-линий (у амфибии) или на границе А- и I-дисков (у млекопитающих) канальцы сети меняют направление и располагаются поперечно, образуя расширенные терминальные или (латеральные) L-цистерны. С поверхности миосимпласта плазмолемма образует длинные трубочки, идущие поперечно в глубину клетки (Т-трубочки) на уровне границ между темными и светлыми дисками. Когда клетка получает сигнал о начале сокращения, этот сигнал перемещается по плазмолемме в виде потенциала действия и распространяется отсюда на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется в цистернах саркоплазматического ретикулума и сокращение миофибрилл прекращается. Для развития усилия сокращения нужна энергия. Она освобождается за счет АТФ- АДФ-превращений. Роль АТФазы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами. Большую роль в деятельности миосимпластов играют включения миоглобина и гликогена. Гликоген служит источником энергии, необходимой не только для совершения мышечной работы, но и поддержания теплового баланса всего организма.Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается из миоглобина и участвует в биохимических реакциях. Миозиновые и актиновые филаменты в быкове почитать, там норм написано
35. Поперечно-полосатая сердечная мышечная ткань. Структурно-функциональная характеристика. Сердечная мышечная ткань Гистогенез и виды клеток. Источники развития сердечной поперечнополосатой мышечной ткани — симметричные участки висцерального листка спланхнотома в шейной части зародыша — так называемые миоэпикардиалъные пластинки. Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 3 вида кардиомиоцитов: 1. рабочие, или типичные, или же сократительные, кардиомиоциты, 2. атипичные кардиомиоциты (сюда входят пейсмекерные, проводящие и переходные кардиомиоциты, а также 3. секреторные кардиомиоциты. Рабочие (сократительные) кардиомиоциты образуют свои цепочки. Укорачиваясь, они обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу. Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Они воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности. Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние — проводящим. Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее — другим проводящим кардиомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим. Секреторные кардиомиоциты выполняют особую функцию. Они вырабатывают гормон - натрийуретический фактор, участвующий в процессах регуляции мочеобразования и в некоторых других процессах. Сократительные кардиомиоциты имеют удлиненную (100—150 мкм) форму, близкую к цилиндрической. Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна (толщиной до 20 мкм). В области контактов клеток образуются так называемые вставочные диски. Кардиомиоциты могут ветвиться и образуют трехмерную сеть. Их поверхности покрыты базальной мембраной, в которую снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро кардиомиоцита (иногда их два) овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения. Миофибриллы слабо обособлены друг от друга, могут расщепляться. Их строение аналогично строению миофибрилл миосимпласта скелетного мышечного волокна. От поверхности плазмолеммы в глубь кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-линии. Их мембраны сближены, контактируют с мембранами гладкой эндоплазматической (т.е. саркоплазматической) сети. Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные утолщения (L-системы), формирующие вместе с Т-трубочками триады или диады. В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта. Кардиомиоциты соединяются друг с другом своими торцевыми концами. Здесь образуются так называемые вставочные диски: эти участки выглядят как тонкие пластинки при увеличении светового микроскопа. Фактически же концы кардиомиоцитов имеют неровную поверхность, поэтому выступы одной клетки входят во впадины другой. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмосомами. К каждой десмосоме со стороны цитоплазмы подходит миофибрилла, закрепляющаяся концом в десмоплакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами (или щелевыми соединениями). Это создает между ними метаболические связи и обеспечивает синхронность сокращений. Возможности регенерации сердечной мышечной ткани. При длительной усиленной работе (например, в условиях постоянно повышенного артериального давления крови) происходит рабочая гипертрофия кардиомиоцитов. Стволовых клеток или клеток-предшественников в сердечной мышечной ткани не обнаружено, поэтому погибающие кардиомиоциты (в частности, при инфаркте миокарда) не восстанавливаются, а замещаются элементами соединительной ткани. 36. Общая характеристика гладкой мышечной ткани . Типы гладких миоцитов ( миоэпителиальные клетки, мионейральные клетки, эндокринные гладкие миоциты, миофибробласты). Строение, локализация и функции. Гладкие мышечные ткани По происхождению различают три группы гладких (или неисчерченных) мышечных тканей — мезенхимные, эпидермальные и нейральные. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2024 stydopedia.ru Все материалы защищены законодательством РФ.
|
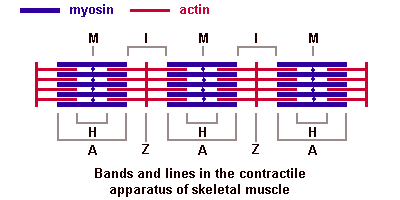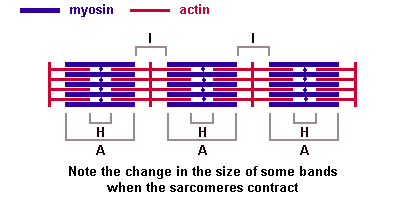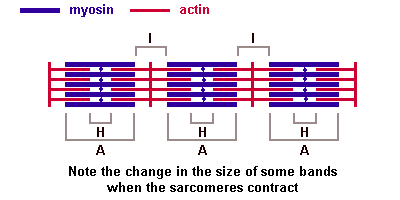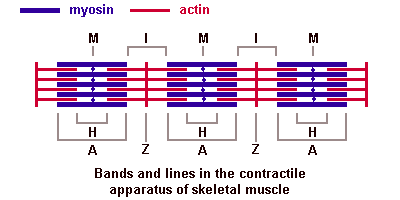 Молекулы миозина имеют длинный хвост и на его конце две головки. При повышении концентрации ионов кальция в области присоединения головок (в своеобразном шарнирном участке) молекула миозина изменяет свою конфигурацию. При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков — тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается.
Молекулы миозина имеют длинный хвост и на его конце две головки. При повышении концентрации ионов кальция в области присоединения головок (в своеобразном шарнирном участке) молекула миозина изменяет свою конфигурацию. При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков — тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается.