|
|
Цитоплазма.Химический состав,физические свойства.Цитоплазма. - обязательная часть клетки, заключенная между плазической мембраной и ядром и представляющая собой вязкое бесцветное основное вещество цитоплазмы, органоиды — постоянные компоненты цитоплазмы и включения — временные компоненты цитоплазмы. Химический состав цитоплазмы разнообразен. Ее основу составляет вода (60—50% всей массы цитоплазмы). Цитоплазма богата белками, в состав цитоплазмы могут входить жиры и жироподобные вещества, различные органические и неорганические соединения. Цитоплазма имеет щелочную реакцию. Одна из характерных особенностей цитоплазмы —постоянное движение (циклоз). Оно обнаруживается прежде всего по перемещению органелл клетки, например хлоропластов.Если движение цитоплазмы прекращается, клетка погибает , так как только находясь в постоянном движении она может выполнять свои функции. Основное вещество цитоплазмы — гиалоплазма (цитозоль) — представляет собой бесцветный, слизистый, густой и прозрачный коллоидный раствор. Именно в нем протекают все процессы обмена веществ, она обеспечивает взаимосвязь ядра и всех органоидов. В зависимости от преобладания в гиалоилазме жидкой части или крупных молекул различают две формы гиалоплазмы: золь — более жидкая гиалоплазма и гель — более густая гиалоплазма. Между ними возможны взаимо переходы: гель легко превращается в золь и наоборот.
Функции гиалоплазмы: Объединение всех компонентов клетки в единую среду Среда для прохождения химических реакций Среда для существования и функционирования органоидов. Гиалоплазма и эргастоплазма.Ультраструктура и функциональное значение. Гиалоплазма (от греч. hyalos — стекло и плазма), основная плазма, матрикс цитоплазмы, сложная бесцветная коллоидная система в клетке, способная к обратимым переходам из золя в гель. В состав Г. входят растворимые белки (ферменты гликолиза, активации аминокислот при биосинтезе белка, многие АТФ-азы и др.), растворимые РНК, полисахариды, липиды. Через Г. идёт транспорт аминокислот, жирных к-т, нуклеотидов, Сахаров, неорганич. ионов, перенос АТФ. Состав Г. определяет буферные и осмотич. свойства клетки.
Цитоплазма эукариотических клеток состоит из полужидкого содержимого и органелл. Основное полужидкое вещество цитоплазмы называют гиалоплазмой (от греч. hyalos — стекло) или матриксом. Гиалоилазма является важной частью клетки, ее внутренней средой.
Она представляет собой сложную коллоидную систему, которая образована белками, нуклеиновыми кислотами, углеводами, водой и другими веществами. В гиалоплазме в растворенном состоянии содержится большое количество аминокислот, нуклео-тидов и других строительных блоков биополимеров, множество промежуточных продуктов, возникающих при синтезе и распаде макромолекул, а также ионов неорганических соединений, таких как Na-, К-, Са2+ Mg2- , Сl-, НС03 , НР042 и др.
Несмотря на то что в электронном микроскопе гиалоплазма выглядит гомогенным веществом, она не является однородной. Гиалоплазма систоит из двух фаз — жидкой и твердой. Жидкая фаза представляет собой коллоидный раствор различных белков и других веществ. В жидкой фазе содержится система тонких (- 2 нм толщиной) белковых нитей — микротрабекул, пересекающих цитоплазму в различных направлениях; это так называемая микротрабекулярная система (рис. 1.7).
Микротрабекулярная система связывает все внутриклеточные структуры. В местах пересечения или соединения концов микротрабекул располагаются группы рибосом.
С микротрабекулярной системой связаны нитевидные, белковые комплексы, или филаменты (тонкие нити) — микротрубочки и микрофиламенты.
Микротрубочки, микрофиламенты и микротрабекулярная система образуют внутриклеточный цитоплазматический скелет (цитоскелет), который упорядочивает размещение всех структурных компонентов клетки Функции гиалоплазмы следующие: 1)Является внутренней средой клетки, в которой происходят многие химические процессы. 2)Объединяет все клеточные структуры и обеспечивает химическое взаимодействие между ними. 3)Определяет местоположение органелл в клетке. 4)Обеспечивает внутриклеточный транспорт веществ и перемещение органелл (например, движение хлоропластов в растительных клетках). 5)Является основным вместилищем и зоной перемещения молекул АТФ. 6)Определяет форму клетки. Эргастоплазмаучастки цитоплазмы клеток растений и животных участки, богатые рибонуклеиновой кислотой (например, глыбки Берга в клетках печени, тельца Ниссля в нейронах). В электронном микроскопе эти участки наблюдаются как упорядоченно расположенные элементы гранулярной эндоплазматической сети. Плазматическая мембрана, её функции.Современные представления о плазматической мембране. Кле́точная мембра́на (или цитолемма, или плазмалемма, или плазматическая мембрана) отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулируют обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определённые условия среды. Функции барьерная— обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами. транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке оптимального pH и концентрации ионов, которые нужны для работы клеточных ферментов. Частицы, по какой-либо причине неспособные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза. При пассивном транспорте вещества пересекают липидный бислой без затрат энергии по градиенту концентрации путем диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа. Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивает в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+). матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие. механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество. энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки; рецепторная — некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы). Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней. ферментативная — мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты. осуществление генерации и проведения биопотенциалов. С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса. маркировка клетки — на мембране есть антигены, действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены. Согласно современным представлениям центральный слой такой мембраны представляет собой текучий липидный бислой с включениями внутримембранных белков. Полагают, что ассоциированные с мембраной белки являются глобулярными. Некоторые из них расположены на полярной поверхности мембраны или частично погружены в ее монослой как с наружной, так и с внутренней стороны. Это так называемые периферические, функционально ассоциированные с мембраной белки, удерживаемые на ее поверхности при помощи нековалентных связей. Другие, интегральные, белки проходят через всю толщу мембраны, в том числе и через внутренние неполярные ее слои. В интегральных белках последовательность аминокислотных остатков распределена таким образом, что гидрофобные остатки аминокислот формируют структуры, которые пронизывают мембрану, а гидрофильные образуют функциональные домены на внутренней и/или наружной поверхности мембраны. Таким образом, функционально разные белки мембраны образуют в жидкокристаллическом бислое фосфолипидов своеобразную мозаичную структуру. Эта мозаика не является строго фиксированной, что позволяет разным классам ФЛ и минорным липидам мембраны при латеральной диффузии формировать определенные кластеры (участки поверхностного монослоя мембраны).
Плазматическая мембрана содержит много гликолипидов, полярные углеводные части которых (остатки моно- и олигосахаридов) расположены на ее поверхности, что позволяет им выполнять специфичные функции, такие как рецепция (клеточное узнавание) и иммунохимические реакции. Выступающие из бислоя гидрофильные олигосахаридные участки гликолипидов образуют у эукариотических клеток подобие наружной оболочки – гликокаликса. Определенную роль в стабилизации липидного бислоя играет и слой воды, покрывающий снаружи монослой фосфолипидов и мембранных белков. Такие монослои воды удерживаются на поверхности мембраны за счет водородных связей между полярными «головками» ФЛ и молекулами воды [2]. В бислое индивидуальные липидные молекулы могут перемещаться (латеральная диффузия), что обеспечивает мембране жидкостность и гибкость. Отдельные молекулы ФЛ в зависимости от длины их жирнокислотных цепей способны перемещаться между наружным и внутренним монослоем мембраны, используя механизм флип-флопа. Все это указывает на то, что бислойная мембрана является единой динамичной и саморегулирующейся системой Модели БМ. 1.4 Эволюция представлений о строении мембран Наличие мембран вокруг живых клеток было установлено более ста лет назад в работах Негели К., который в 1855 г. обнаружил, что неповрежденные клетки могут изменять свой объем при изменении осмотического давления окружающей среды. Эти исследования были продолжены Овертоном Е., показавшим, что неполярные молекулы легче проходят через клеточную мембрану, чем полярные соединения. На основе этих наблюдений он впервые высказал предположение, что клеточная мембрана имеет липидную природу. Развитие идей о структуре мембран существенно продвинулось благодаря работам Гортера Е. и Грендела Ф., проведенным в 1925 г. Эти авторы впервые выдвинули концепцию липидного бислоя. Идея возникла на основе простого эксперимента. Липиды эритроцитов экстрагировали ацетоном и затем получали из них тонкую пленку на поверхности воды. С помощью поплавка сжимали слой липидных молекул на границе раздела вода–воздух до тех пор, пока этот слой не начинал оказывать сопротивление дальнейшему сжатию; это явление было объяснено образованием плотно упакованной мономолекулярной липидной пленки. Измерение площади, занимаемой липидами, и сравнение ее с площадью поверхности эритроцитов, из которых эти липиды были экстрагированы, дали соотношение 2:1. Отсюда был сделан вывод, что мембрана эритроцитов состоит из липидных молекул, расположенных в два слоя. По-видимому, этот вывод Гортера Е. и Грендела Ф. оказался правильным только благодаря взаимной компенсации ошибок (во-первых, экстракция ацетоном извлекает не все липиды, во-вторых, они дали заниженную оценку площади поверхности эритроцитов, использовав для ее определения высушенные клетки). Однако в историческом плане эта работа имела большое значение, поскольку концепция липидного бислоя как структурной основы биологических мембран на самом деле оказалась верной. Мысль о том, что с мембранами связаны белки, высказана десятью годами позже Даниелли Дж. в связи с необходимостью объяснить явное расхождение между поверхностным натяжением на границах раздела масло–вода и мембрана–вода. Была высказана гипотеза, что мембрана состоит из двойного липидного слоя, и предположено, что белок располагается на ее поверхности – модель Даниелли–Дэвисона, или модель «сэндвича» (рисунок 1.2). 1 – углеводородные гидрофобные цепочки; 2 – полярные гидрофильные группы молекулы; 3 – полярные поры, по которым вещества диффундируют в клетку Рисунок 1.2 – Модель строения биологических мембран Даниелли–Девисона На рисунке 1.2 показан бимолекулярный липидный слой, окруженный с двух сторон монослоями белка. Это была очень удачная модель, и в течение последующих 30 лет многочисленные экспериментальные данные, особенно полученные с помощью дифракции рентгеновских лучей и электронной микроскопии, полностью подтвердили ее адекватность. Основными компонентами биологической мембраны являются липид и белок, вопрос о взаимном расположении этих компонентов в мембране стал предметом многочисленных дискуссий, так как обнаружилось, что мембраны выполняют разнообразные функции.
В 1959 г. Робертсон Дж. Д. предположил, что все клеточные мембраны построены по единому принципу, и высказал концепцию унитарной (или единообразной) мембраны (рисунок 1.3).
Рисунок 1.3 – Унитарная схема асимметричного строения биомембраны Робертсона. Предложенная модель во многом сходна с классической моделью Даниелли Дж.: основу мембраны составляет липидный бислой, а нелипидные компоненты (прежде всего белки) в полностью развернутой конформации лежат на поверхности бислоя, связываясь с липидами за счет электростатических и гидрофобных взаимодействий. В модели Робертсона нашла отражение еще одна важная структурная особенность мембраны – ее асимметрия. Последующий прогресс в мембранологии, в результате которого сформировались современные представления о структуре биомембран, в значительной мере был достигнут благодаря успехам в изучении свойств мембранных белков. Электронно-микроскопические исследования с применением метода замораживания–скалывания показали, что в мембраны встроены глобулярные частицы, а биохимикам с помощью детергентов удалось диссоциировать мембраны до состояния функционально активных «частиц». Данные спектральных исследований указывали, что для мембранных белков характерно высокое содержание α-спиралей и что они, вероятно, образуют глобулы, а не распределены в виде монослоя на поверхности липидного бислоя. Неполярные свойства мембранных белков наводили на мысль о наличии гидрофобных контактов между белками и внутренней неполярной областью липидного бислоя. Тогда же были разработаны методы, позволившие выявить текучесть липидного бислоя. Сингер и Николсон свели воедино все эти идеи, предложив в 1972 г. новую модель молекулярной организации биомембран – жидкостно-мозаичную модель (рисунок 1.4).
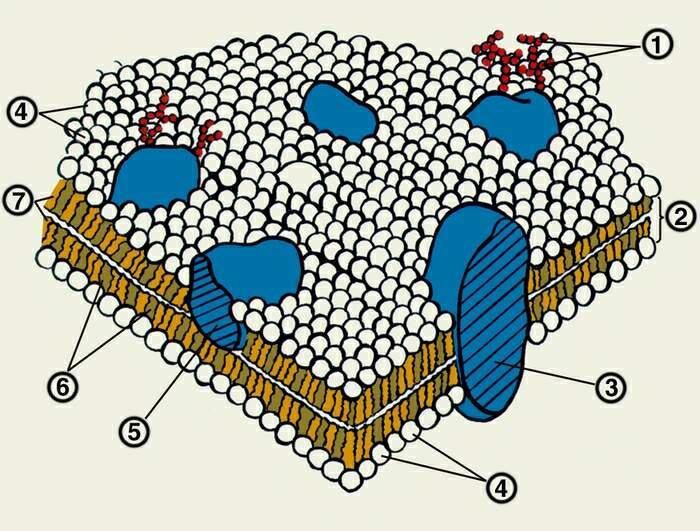
1 – углеводные фрагменты гликопротеидов; 2 – липидный бислой; 3 – интегральный белок; 4 – «головки» фосфолипидов; 5 – периферический белок; 6 – холестерин; 7 – жирнокислотные «хвосты» фосфолипидов. Рисунок 1.4 – Модель жидкостно-мозаичной мембраны Сингера и Николсона Согласно жидкостно-мозаичной модели: 1) Структурной основой биомембран является липидный бислой, в котором углеводородные цепи молекул фосфолипидов находятся в жидкокристаллическом состоянии. 2) В липидный бислой, имеющий вязкость растительного масла, погружены или встроены молекулы белков, способные передвигаться по мембране. В противоположность прежним моделям, рассматривающим мембраны как системы из жестко фиксированных компонентов, жидкостно-мозаичная модель представляет мембрану, как «море» жидких липидов, в котором плавают «айсберги» белков. В зависимости от прочности связи с мембраной белки в рамках мозаичной модели подразделяются на два типа: периферические и интегральные. К периферическим относятся белки, которые связаны с мембраной за счет полярных и ионных взаимодействий и относительно легко отделяются от нее в мягких условиях, например, при промывании буферными растворами с различными значениями рН или ионной силы либо растворами, содержащими комплексообразующие вещества типа ЭДТА. Интегральные белки имеют на своей поверхности большие гидрофобные участки и располагаются внутри мембраны. Для выделения интегральных белков необходимо сначала разрушить липидный бислой. Жидкостно-мозаичная модель строения биомембран в настоящее время является общепризнанной, однако следует помнить, что она все же представляет собой упрощенное и схематичное отражение такой сложной и разносторонней системы, как биологическая мембрана. Одним из постулатов этой модели является предположение о свободном движении молекул белков и липидов в двумерной фазе липидного бислоя. Однако вскоре выяснилось, что не все белки и липиды способны к свободному перемещению, в некоторых случаях их подвижность сильно ограничена. Во многих мембранах интегральные белки находятся в фиксированных положениях за счет высокой концентрации белка вследствие его агрегации, образования липидных доменов, а также взаимодействия белков с цитоскелетом, образуемым внутренними структурами клетки. В некоторых мембранах значительные количества липидов могут находиться в сильно упорядоченном состоянии или, наоборот, в составе небислойных фаз. Это означает, что распределение липидов вдоль поверхности мембраны не является гомогенным, как следовало бы ожидать в случае их свободной диффузии согласно жидкостно-мозаичной модели, а в значительной мере гетерогенно [1]. Кроме того, жидкостно-мозаичная модель не объясняет высокую гетерогенность липидного состава биологических мембран. Необходимо отметить, что липиды биологических мембран различаются не только по структуре полярных групп, но и по степени ненасыщенности и длине углеводородных цепей, а также по способу их присоединения к глицериновому остатку (сложная эфирная, простая эфирная и винильно-эфирная связь). Липидный состав биологических мембран всегда чрезвычайно гетерогенен, и в его построении участвуют сотни химически индивидуальных липидных молекул. Данный факт не согласуется с представлениями о пассивной роли липидов в функционировании мембран в качестве структурной матрицы, в которой расположены мембранные белки [1]. Несмотря на это в настоящее время по-прежнему пользуются жидкостно-мозаичной моделью строения мембраны, но в усложненной форме, в которой отражены новые, специфические, не известные ранее закономерности. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2024 stydopedia.ru Все материалы защищены законодательством РФ.
|